Paleogenòmica: Els laboratoris d’arqueogenètica de Pontus Skoglund i de Linus Girland Flink han aconseguit la reconstrucció del genoma complet d’un individu que va viure en el període de l’Imperi Antic d’Egipte, tal com expliquen en un article publicat a Nature, amb Adeline Morez Jacobs com a primera autora. Cal recordar que fins ara hom no havia aconseguit el genoma complet de cap ésser humà que hagués viscut a l’antic Egipte, és a dir a l’Egipte del període comprès entre el 3150 a.C. i l’any 30 a.C. Les condicions climàtiques fan poc avinent la preservació de l’ADN. Jacobs et al. pensen que genomes complets d’aquesta civilització poden ajudar a entendre la interconnectivitat regional de l’època. En aquest article es presenten un genoma complet amb un grau de cobertura de 2× corresponent a un home adult soterrat a Nuwayrat (نويرات), a l’Alt Egipte. El radiocarboni indica que aquesta tomba seria del període 2855-2570 a.C., és a dir uns pocs segles després de la unificació de l’Alt i del Baix Egipte, corresponent a allò que els historiadors denominen primer període dinàstic i regne antic d’Egipte. Aquest cos fou soterrat en un pot ceràmic en una tomba tallada a la roca, la qual cosa hauria ajudat a la preservació d’ADN. La majoria del seu genoma es correspon a una ascendència neolítica nord-africana, però vora un 20% es correspondria a l’orient del Creixent Fèrtil, és a dir Mesopotàmia i regions veïnes. Aquesta afinitat genètica és semblant a l’observada en genomes d’Anatòlia i Pròxim Orient del Neolític i de l’Edat de Bronze. Jacobs et al. són conscients que cal conèixer més genomes per entendre la diversitat dels primers egipcis, però les dades d’aquest ja indiquen que els contactes entre Egipte i Mesopotàmia no es limitaren a objectes i imatges (dels quals hi ha testimoni arqueològic en forma d’animals i plantes domèstics, sistemes d’escriptura, etc.) sinó també una forma de migració humana, si més no des de Mesopotàmia a Egipte.
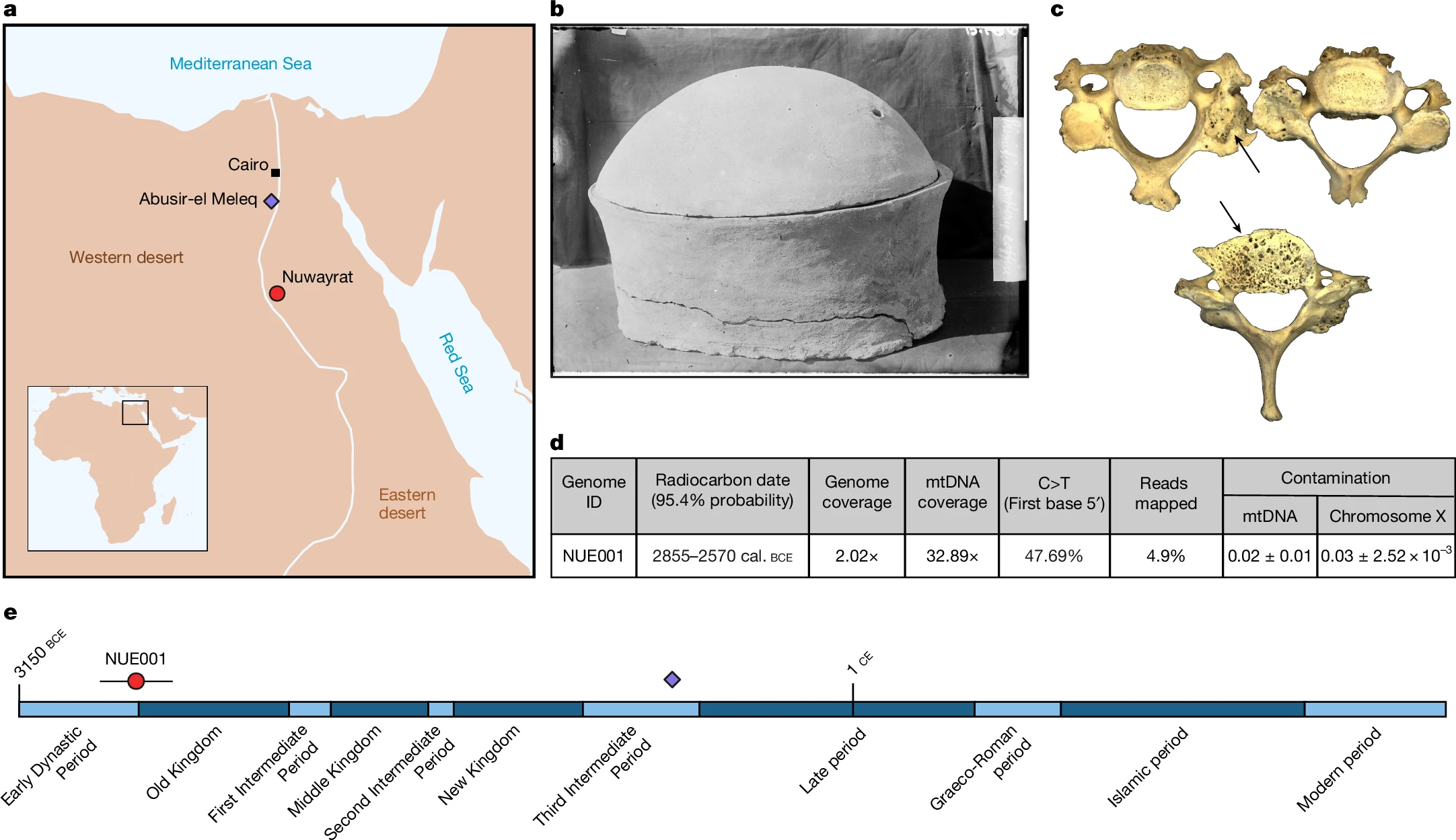
El genoma antic NUE001 es correspon a un individu que va rebre sepultura a Nuwayrat dins d’un pot de ceràmica. Les dades de radiocarboni indiquen que va viure en algun moment del període del 2855-2570 a.C.
La paleogenòmica de l’Antic Egipte
Els egiptòlegs ens diuen que la civilització dinàstica d’Egipte romangué durant milers d’anys, aproximadament entre el 3150 a.C. (sorgiment de la primera dinastia) fins el 30 a.C. (final de la dinastia ptolemaica i incorporació a l’Imperi Romà). L’Antic Egipte ens impressiona per l’arquitectura monumental, per la tecnologia sofisticada, per l’estabilitat relativa dels sistemes de creences, etc. Tenim la impressió que és la civilització humana que més ha perdurat en el temps: més de tres mil·lennis.
La Dinastia I d’Egipte es defineix per l’assoliment de la unificació política del Baix Egipte (nord) i de l’Alt Egipte (sud), que tingué lloc a final del IV mil·lenni a.C. L’Antic Regne d’Egipte es defineix com el període situat entre els anys 2686 a.C. i 2125 a.C. Durant aquests cinc segles llargs, Egipte conegué considerables avenços, reflectits en la construcció del primer complex de piràmide esglaonada del rei Djoser, i en la Gran Piràmide de Giza del rei Khufu. Hom ha assumit que la població de l’Antic Regne d’Egipte era essencialment local o autòctona, amb una aportació limitada de les regions veïnes. Ara bé, les dades arqueològiques indiquen que hi havia importants connexions comercials dins del marc del Creixent Fèrtil, és a dir els països que envolten i inclouen Egipte i Mesopotàmia, ja des del VI mil·lenni a.C. Possiblement ja abans, durant l’anomenada revolució neolítica (domesticació d’animals i plantes), s’articulà aquest espai. A final del IV mil·lenni a.C., amb el creixement de la civilització sumèria a Mesopotàmia el bescanvi cultural continuà a desenvolupar-se. És en aquest darrer període on fan aparició a Egipte innovacions com el torn ceràmic. Les primeres evidències d’escriptura jeroglífica es troben en peces d’ivori de la tomba U-j d’Abidos, datades entre el 3320 i el 3150 a.C.
La bioarqueologia té molt a dir-nos sobre l’antic Egipte. Els estudis de morfologia dental han remarcat la relació dels antics egipcis amb les poblacions de l’Àfrica del Nord i de l’Àsia Occidental. En termes de paleogenòmica, però, no hi ha hagut gaire fortuna. Les primeres tecnologies d’ADN antic foren assajades en restes de l’antic Egipte, però sense èxit. Hom assumí que les condicions climàtiques regionals eren enemigues de la preservació de l’ADN. Tot i amb tot, en el 2017 es publicaren dades d’ADN nuclear de tres individus d’Abusir el-Meleq, que haurien viscut en el període tardà (787-23 a.C.). Ara bé, no es tracta d’un genoma complet, sinó més aviat d’un genotipatge de 90.000 a 400.000 dianes genètiques.
En imaginar-nos l’Antic Egipte som en deute encara amb les escoles històriques del període tardà, que coneixem bàsicament per les referències que Flavi Josep fa de l’historiador egipci Manetó. D’ells derivem la llista d’una trentena de dinasties. Els egiptòlegs moderns les han agrupades en un esquema de períodes estables i de períodes inestables o intermedis. Així, hom parla de tres períodes intermedis, marcats per guerres de gran abast, per l’ocupació de governants estrangers i/o per col·lapse polític intern. Cal pensar que en aquests períodes intermedis es visqueren alteracions de l’estructura genètica de la població egípcia. Per això és valuós disposar del genoma complet anterior al ‘primer període intermedi’, que ens presenten Jacobs et al.
Es tracta d’una seqüència genòmica completa amb un grau de cobertura de 2,02, obtinguda de restes d’un individu recuperar d’una necròpolis de Nuerat (Nuwayrat).
L’individu NUE001
Nuwayrat se situa prop de la localitat de Beni Hasan, a uns 265 km al sud de l’actual capital, Cairo. La datació per radiocarboni indica que les restes esquelètiques NUE001 procediren de l’interval 2855-2570 a.C., amb una probabilitat del 95,4%. Aquesta forquilla cobreix part del període Dinàstic Inicial i del període del Regne Antic. Els arqueòlegs, atenent a la cultura material i a les pràctiques funeràries d’aquesta necròpoli, s’inclinen per una datació entre la III i la IV Dinastia, ja de ple en el Regne Antic.
El cos NUE001 fou dipositat en un gran vas ceràmic dins d’una tomba tallada a la roca. Observacions en els cementiris reials del Primer Període Dinàstic i del Regne Antic de prop de la ciutat de Memfis, conviden a pensar que aquest tractament funerari era reservat habitualment a individus de classe social alta.
Les dades genòmiques de NUE001 indicarien uns possibles trets fenotípics, amb tota la reserva que calgui per la manca d’estudi d’aquesta població: ulls bruns, cabells bruns, pell fosca o negra. La dotació cromosòmica sexual era XY, coincident amb un trets esquelètics masculins. L’examen osteològic indicaria una alçada d’entre 157,4 i 160,5 cm. Va morir a una edat d’entre 44 i 64 anys, més probablement cap a aquest darrer extrem, cosa que es pot considerar una edat avançada per l’època. Aquesta edat avançada es manifesta en unes dents gastades, en osteoartritis en moltes articulacions i vèrtebres. Indicadors d’estrès suggereixen que va viure sota un període extens de treball físic, cosa que contrasta amb el tractament funerari referit. Podria haver estat un terrissaire.
Les dades genòmiques lliguen bé amb els trets morfològics dentals i les mesures cranianes.
En l’esmalt i en la col·làgena del segon molar inferior esquerra s’han fet anàlisis multi-isotòpiques (δ13C, δ15N, δ18O and 87Sr/86Sr). Això pot ajudar a entendre la dieta que feia de petit o el seu origen geogràfic. Les dades isotòpiques indicarien que va créixer en el clima càlid i eixut de la Vall del Nil. En la infantesa seguí una alimentació omnívora, basada en proteïnes d’animals terrestres i de plantes (blat, ordi). Aquesta dieta era la típica de l’Antic Egipte fins al període còptic. Els valors de δ15N suggereixen un consum d’aliments produïts en camps adobats i/o la inclusió de peixos del Nil.
La seqüenciació genòmica
Es prepararen set extractes de ciment enriquit en ADN en genoteques de seqüenciació d’ADN monocatenari, rastrejat en una plataforma Illumina. Cinc d’aquestes genoteques mostraven patrons de degradació típics d’ADN antic: alta taxa de substitucions citosina-a-timina. Les altres dues genoteques fores descartades per indicis de contaminació elevada.
Les genoteques Y11475 i T11477 foren seqüenciades addicionalment en Illumina NovaSeq 6000 i NovaSeq X, obtenint-ne 8300 milions de 2 x 100 parells llegits.
El genoma resultant, NUE001, fou fusionat amb els 3233 genomes d’individus actuals de Human Origins Array i amb 805 d’individus antics. L’anàlisi de components principals indica que NUE001 se situa dins de persones actuals del nord d’Àfrica i de l’oest d’Àsia. NUE001 pertany a l’haplogrup mitocondrial I/N1a1b2 i l’haplogrup del cromosoma Y E1b1b1b2b~: aquests haplogrups són els més comuns en l’actualitat a l’Àfrica del Nord i a l’Àsia de l’Oest. L’examen d’homozigositat sobre regions de més de 4 centimorgans indica que NUE001 no presenta cap consanguinitat recent en la seva ascendència.
L’ascendència de NUE001
El marc qpAdm considera diferents poblacions humanes. Jacobs et al. tenen en compte un conjunt de 13 poblacions, incloent-hi les etapes neolítica i calcolítica de l’Àsia Occidental Neolítica, l’Àfrica del Nord i la Regió Mediterrània. Això resulta en NUE001 en la barreja següent:
- 77,6 ± 3,8% del neolític mitjà nord-africà, representat pel jaciment marroquí de Skhirat-Rouazi del 4780-4230 a.C.
- 22,4 ± 3,8% del neolític mesopotàmic de 9000-8000 a.C.
Altres models indiquen una tercera aportació (4,7 ± 8,2%) del Llevant neolític/calcolític.
Les dades corroboren la continuïtat entre la població neolítica egípcia i la població del Primer Període Dinàstic/Regne Antic que ja ens comunica la cultura material i les anàlisis bioarqueològiques. Ara bé, també hi hauria hagut una aportació atribuïbles a una immigració a Egipte procedent de l’Àsia Occidental. Jacobs et al. consideren que cal encara disposar de més estudis d’ADN antic del període neolític de l’Àfrica del Nord per arribar a hipòtesis més detallades.
Jacobs et al. han estudiat la relació entre NUE001 i els genotips d’Abusir-el Meleq del tercer període intermedi (787-544 a.C.). Tenen en compte ara nou poblacions de l’Àfrica del Nord, Àsia Occidental i Grècia. La continuïtat entre els dos no és pas completa, ja que en els egipcis del Tercer Període Intermedi havien rebut una major aportació asiàtica. Aquesta aportació asiàtica podria relacionar-se amb l’expansió cananea de l’Edat de Bronze, que començaria a final del Regne Mitjà d’Egipte.
Pel que fa a la continuïtat amb les poblacions egípcies actuals, Jacobs et al. consideren que aquestes tenen cinc orígens bàsics:
- (1) Nuwayrat (32,1–74,7%).
- (2) Neolític Mitjà marroquí (28,9–72,7%).
- 3) Llevant de l’Edat de Bronze (11,6–57,1%).
- (4) la població ‘Mota’ de l’Etiòpia de fa 4500 anys (7,4–56,0%).
- (5) dos individus del Congo fa 230 anys (4.8–52.0%).
Lligams:
- Whole-genome ancestry of an Old Kingdom Egyptian. Adeline Morez Jacobs, Joel D. Irish, Ashley Cooke, Kyriaki Anastasiadou, Christopher Barrington, Alexandre Gilardet, Monica Kelly, Marina Silva, Leo Speidel, Frankie Tait, Mia Williams, Nicolas Brucato, Francois-Xavier Ricaut, Caroline Wilkinson, Richard Madgwick, Emily Holt, Alexandra J. Nederbragt, Edward Inglis, Mateja Hajdinjak, Pontus Skoglund & Linus Girdland-Flink. Nature (2025).
