Exòmica: La dreta és la mà directora de la majoria de persones. En els esquerrans ho és la mà esquerra. Aquesta preferència diferencial és una manifestació de l’especialització dels hemisferis del cervell. El percentatge d’esquerrans puja en poblacions diagnosticades de diversos trastorns de desenvolupament neural. Els estudis d’associació genòmica han assenyalat que la mà directora o l’asimetria cerebral s’associen amb variants majoritàriament externes a les regions codificadores de proteïna, que deuen actuar com a reguladores indirectes d’expressió gènica. Entre els gens afectats per aquestes variants hi ha els gens de tubulines i de proteïnes associades a microtúbuls. Els microtúbuls de tubulina participen en el citoesquelet i mecanismes de transport de gairebé tots els tipus cel·lulars, però són especialment rellevants en neurones. Clyde Francks, del Departament de Llenguatge i Genètica de l’Institut Max Planck, ha coordinat una recerca exòmica sobre la lateralitat de la mà directora, que apareix aquesta setmana en forma d’article a Nature Communications, amb Dick Schijven, de la Radboud Universiteit, com a primer autor. Schijven et al. utilitzen dades extretes del UK Biobank que abasten 38.043 persones esquerranes i 313.271 persones dretanes. S’hi fixen en variants codificants rares, és a dir que tenen freqüències gèniques inferiors a l’1%. De tots els gens analitzats, és el gen de la beta-tubulina TUBB4B el que mostra una major associació per a la lateralitat manual. De mitjana la freqüència de variants rares de TUBB4B és 2,7 vegades superior en esquerrans respecte dels dretans. La majoria de variants són modificacions puntuals que es troben en heterozigosi. Hi ha dues variants d’alteració de la pauta de lectura que es troben exclusivament en esquerrans. Val a dir que Schijven et al. no han trobat en el seu estudi variants de TUBB4B que se saben vinculades a malalties sensorineurals i/o ciliopàtiques. Més enllà del gen TUBB4B, Schijven et al. troben associacions amb variants dels gens DSCAM i FOXP1, gens que altres estudis exòmics vinculen a l’autisme o a l’esquizofrènia. Schijven et al. calculen que l’heretabilitat d’ésser esquerrà degut a aquestes variants codificants rares és de tan sols el 0,91%. El més rellevant d’aquesta recerca és que ofereix més indicis sobre el paper dels microtúbuls en l’asimetria cerebral.
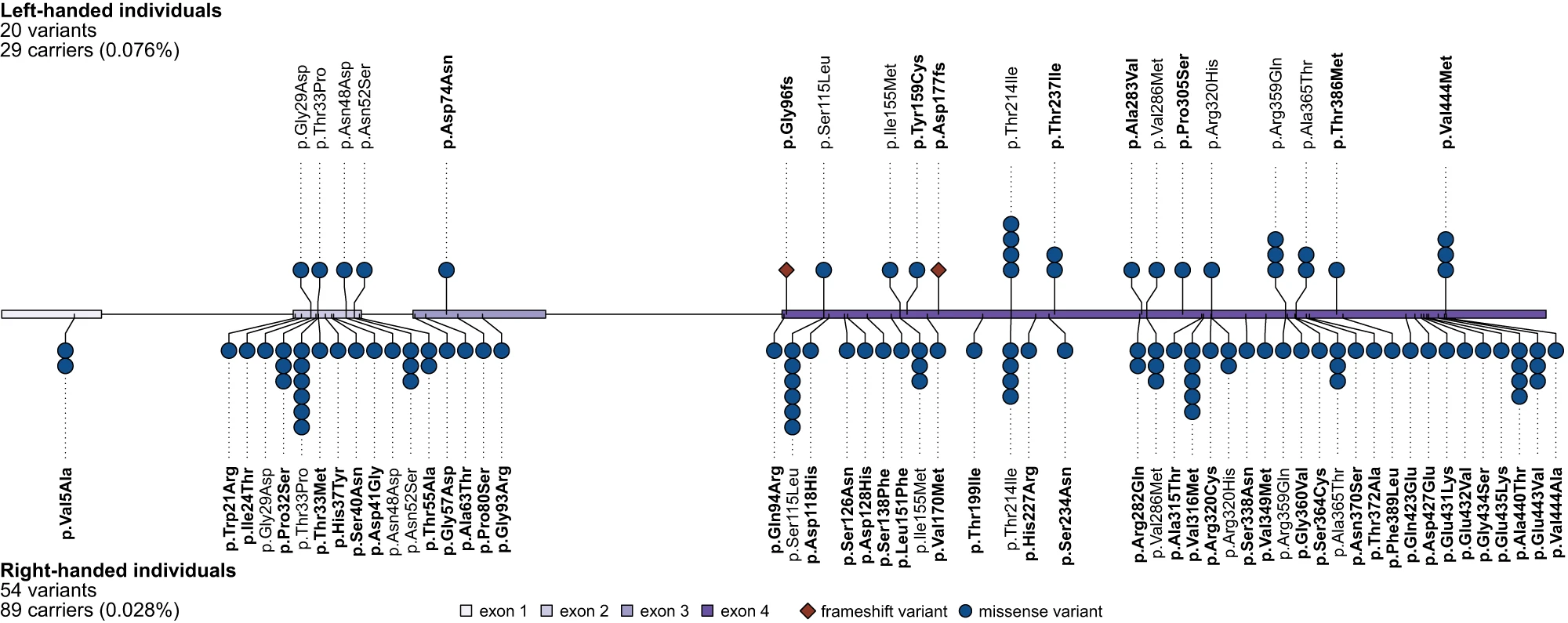
Schijven et al. han trobat un grapat de variants rares del gen TUBB4 en el UK Biobank. Proporcionalment, n'hi ha més entre les persones esquerranes
Els esquerrans: el 10% de la humanitat
Aquesta recerca ha estat concebuda per Dick Schijven, Sourena Soheili-Nezhad (investigador del MPI) i Clyde Francks. Les dades foren tractades per Schijven i Soheili-Nezhad. L’anàlisi formal fou conduït per Schijven. Els fons de recerca foren aportats per Francks i Simon E. Fisher (director del Departament de Llenguatge i Genètica del MPI). L’article fou tramès a Nature Communications el 21 de juliol del 2023, i fou acceptat després d’un procés de revisió el 20 de febrer.
Vora el 90% de la població humana és dretana, i vora el 10% és esquerrana. Encara que aquesta proporció pot variar en l’espai i en el temps (del 2% al 14%), és prou consistent la majoria dretana. La lateralitat manual seria una manifestació de l’asimetria cerebral, de manera que en els dretans hi ha un domini de l’hemisferi cerebral esquerre, i en els esquerrans de l’hemisferi dret. La lateralitat cerebral també es manifesta en el fet que les regions i xarxes importants per a la llengua es concentren en l’hemisferi esquerre per a la majoria de persones.
L’asimetria cerebral apareix aviat en el desenvolupament, i ja es registra en els moviments de braços a les 10 setmanes d’edat gestacional. Així doncs, respon a un programa regulat genèticament en l’eix esquerra-dreta del sistema nerviós central.
Els estudis de bessons indiquen que l’heretabilitat d’ésser esquerrà és del 25%. Ara bé, els estudis d’associació genòmica realitzats en mostres de menys de 10.000 individus no han estat capaços d’identificar gens concrets. Estudis més grans, basats en el UK Biobank (amb 30.000 esquerrans i 300.000 dretans), han trobat tres o quatre loci genòmics amb associació significativa amb ésser esquerrà. Aquests gens són TUBB (gen de la beta-tubilina), MAP2 i MAPT (proteïnes associades a microtúbuls). Les tubulines són les proteïnes que formen la paret dels microtúbuls, els quals són un dels elements del citoesquelet, i participen en processos de creixement, divisió cel·lular, migració, morfologia, formació d’eixos, creixement d’axons, transport intracel·lular, etc. És possible que els microtúbuls contribueixen a la quiralitat cel·lular en el cervell i així a la formació d’una asimetria esquerra-dreta.
Esquerrans i dretans formen part de la variació normal de la població humana. No obstant, en esquerrans hi ha una major incidència d’autisme i d’esquizofrènia. Hom ha pensat que això es deuria a variants genètiques rares, les quals contribuirien en una part relativament petita a la lateralitat esquerrana.
Schijven et al. utilitzen dades de seqüències exòmiques (els exomes són la fracció del genoma d’ADN que es transcriu a ARN missatger i que es tradueix a proteïna) del UK Biobank per investigar la contribució de variants codificants rares a l’esquerranitat.
313.271 dretans i 38.043 esquerrans
Aquesta és la base de dades del UK Biobank que empren Schijven et al. El UK Biobank inclou població britànica. Schijven et al. classifiquen aquestes persones en 1) origen asiàtic; 2) origen negre; 3) origen xinès; i 4) origen blanc. La taxa d’esquerrans varien en aquests quatre grups, probablement perquè en algunes cultures es força l’ús de la mà dreta com a mà directora. Schijven et al. opten per realitzar l’estudi d’associació de variants genètiques amb la lateralitat manual per a cadascun dels quatre grups d’ascendència.
Schijven et al. exclouen de l’anàlisi 6.511 individus del UK Biobank que reportaven un ús equivalent de les seves dues mans.
L’associació es fixa en variants exòniques amb freqüències gèniques rares (≤1%), sempre que siguin seqüències de qualitat i tinguin una probabilitat alta d’afectar la funció proteica. Això inclou aquelles variants que alteren la pauta de lectura o que introdueixen codons de stop més enllà del 5% terminal. També inclou les variants que comporten un canvi en la cadena peptídica que sigui prou rellevant sobre la predicció funcional.
Schijven et al. analitzen 18.381 gens seguint un criteri estricte de variants, que puja a 18.925 gens quan es relaxa aquest criteri.
De tots aquests gens tan sols un, TUBB4B mostra una associació significativa amb la lateralitat manual. Aquest gen codifica per a un dels components dels microtúbuls, la beta-tubulina. Amb associacions menys marcades apareixen el gen TRAK1 (implicat en el tràfic mitocondrial en axons) i el gen MTMR6 (que codifica una fosfatasa de miotubularina).
Schijven examinen els resultats de 48 gens que estudis d’associació genòmica han vinculat amb l’esquerranitat. D’aquests el que apareix amb més força en l’estudi de variants exòniques rares és FOXN2, que codifica per un factor de transcripció de cèl·lules epitelials i del sistema nerviós embrionari.
Variants rares de TUBB4B
Schijven et al. han observat 20 variants rares de TUBB4B en 29 esquerrans, i 53 variants rares en 89 dretans. Això significa que la variants rares de TUBB4B són 2,7 vegades més freqüents en esquerrans que en dretans.
Totes aquestes variants es troben en heterozigosi. Només en un dels individus s’acumulen dues variants (però totes dues en heteorozigosi).
La majoria de les variants seleccionades de TUBB4B produeixen la substitució d’un aminoàcid per un altre en la seqüència proteica. Hi ha dues variants en persones esquerranes que indueixen una alteració en la pauta de lectura (no n’hi ha cap en dretans): aquesta alteració de la pauta de lectura condueix molt probablement a un al·lel no-funcional i, per tant, a una situació d’haploinsuficiència.
Cal remarcar que la seqüència aminoacídica de TUBB4B és molt conservadora. L’homologia entre tetràpodes no baixa del 99,3%, i és superior al 99,8% entre mamífers. Val a dir que la beta-tubulina també es codificada per altres vuit gens en el genoma humà: TUBB, TUBB1, TUBB2A, TUBB2B, TUBB3, TUBB4A, TUBB6 i TUBB8.
Schijven han examinat si les variants rares de TUBB4B causen en els portadors alteracions en la parla, en la visió, en l’audició, etc., i no hi troben res. Això s’explica si recordem que són variants en heterozigosi, i que la beta-tubulina compta amb vuit còpies en el genoma haploide.
La regressió de la càrrega d’heretabilitat d’aquestes variants sobre l’esquerranitat indica que és del 0,91%.
Portadors de variants rares de TUBB4B, de DSCAM i FOXP1 tenen una major probabilitat d’ésser esquerrans. Val a dir, però, que la immensa majoria d’esquerrans no són portadors d’aquestes variants. Hi ha una munió de factors que determinen l’asimetria cerebral i la lateralitat manual, però els microtúbuls semblen tenir un paper especial. Aquestes variants rares deuen ser al·lels dominants, capaços d’alterar en heterozigosi la quiralitat de les cèl·lules del sistema nerviós embrionari. Els microtúbuls actuarien sobre la quiralitat cel·lular per la seva implicació en l’estructura i funcionament dels undulipodis, que no tan sols participen en la motilitat cel·lular sinó també en la creació de corrents en el medi extracel·lular.
El grup de recerca de Francks és expert en neuroimatge, però en el UK Biobank només hi ha dades de MRI del cervell per a 13 dels portadors de variants rares de TUBB4B. Disposar de bases de dades més àmplies ajudarà a entendre millor el paper dels microtúbuls en l’asimetria cerebral.
Lligams:
- Exome-wide analysis implicates rare protein-altering variants in human handedness. Dick Schijven, Sourena Soheili-Nezhad, Simon E. Fisher & Clyde Francks. Nature Communications 15: 2632 (2024).
- Plataforma d’Anàlisis de Recerca del UK Biobank.


