Selenologia: L’Observatori Estratosfèric d’Astronomia d’Infraroig (SOFIA) és un Boeing 747SP equipat amb un telescopi reflector de 2,7 metres d’obertura, que vola a una altitud de 12 a 14 km. SOFIA és el resultat d’una col·laboració entre la NASA i el Deutsches Zentrum für Luft- und Raumfahrt (DLR). Fou amb el SOFIA que Casey I. Honniball realitzà la seva tesi doctoral sobre la detecció remota de components volàtils de la Terra i de la Lluna, que defensà reeixidament l’octubre del 2019. Avui la revista Nature Astronomy publica un article encapçalat per Honniball on reporta la detecció, en el marc d’aquests treballs, d’aigua molecular en la superfície lunar directament sotmesa a un cicle de dia:nit. Estudis anteriors havien detectat un tret d’absorció característic a una longitud d’ona de 3 μm, que s’interpreta com a signe d’una hidratació generalitzada de la superfície lunar. Aquest senyal, però, no discrimina si la hidració és deguda a aigua molecular (H2O) o altres compostos hidroxílics (OH). Per discriminar-ho hom pot fer ús d’una signatura espectral a 6 μm, que és específica de l’H2O, i això és el que han fet Honniball et al. amb SOFIA. A latituds lunars altes hom troba efectivament aquest senyal. L’abundància en aquestes regions seria de 100-400 ppm (en termes de massa). Certament el rang de latitud del senyal és estret i, a més, la distribució s’associa estretament amb la geologia local, de forma que seria pas un fenomen global. A més, la major part de l’aigua detectada es trobaria en l’interior de materials vitris o en cavitats entre grans que queden protegides de l’ambient superficial. Altrament, aquesta aigua no podria romandre en la superfície del nostre planeta satèl·lit.
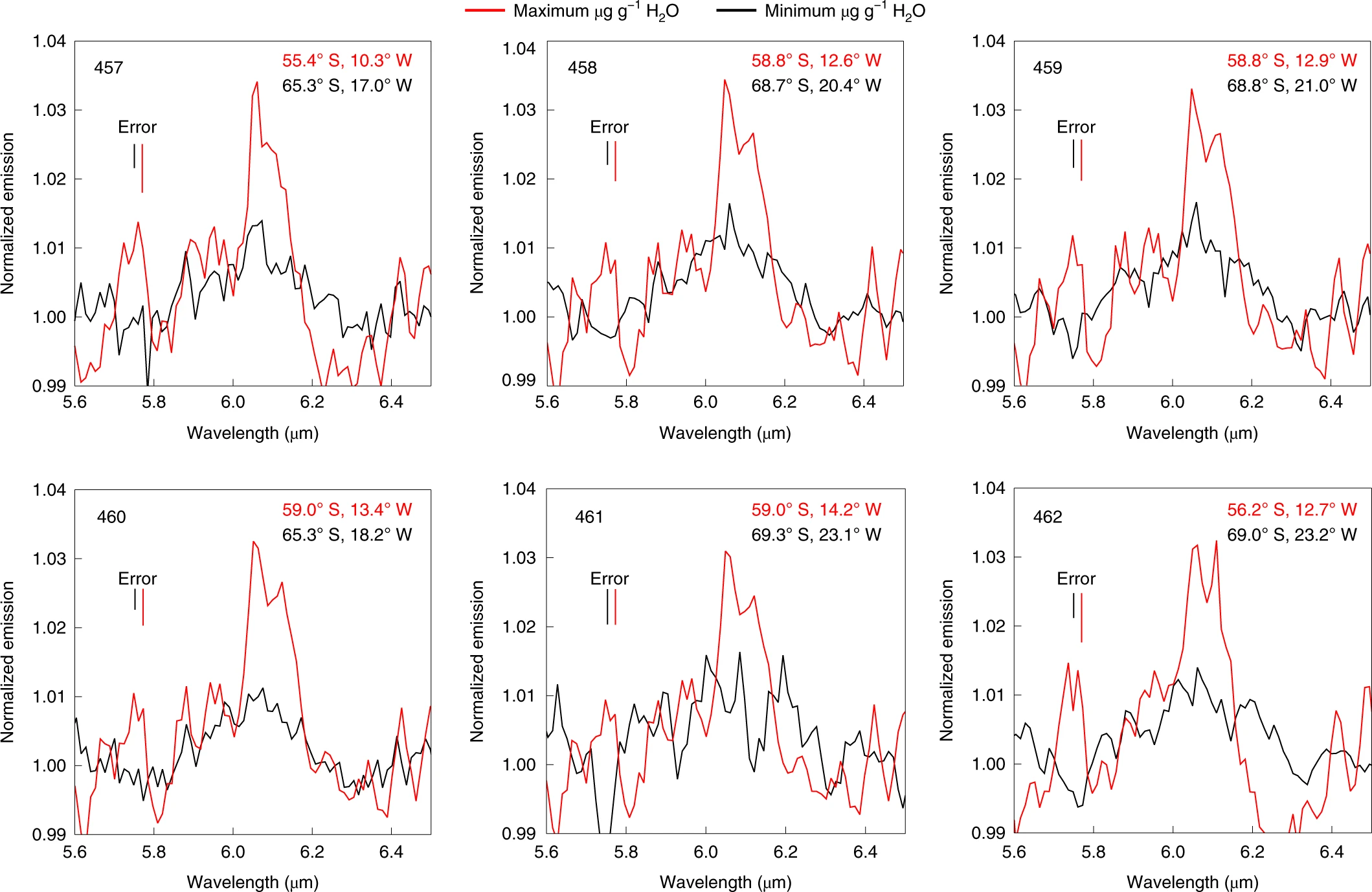
Espectres d’infraroig obtinguts de la regió del cràter Clavius. El pic corresponent a una longitud d’ona de 6000 nm es correspondria a l’H2O
Observacions del 31 d’agost del 2018
Les observacions en aquest estudi foren realitzades per Honniball i Paul G. Lucey, del Hawaiʻi Institute of Geophysics and Planetology, de la University of Hawaiʻi at Mānoa, de Honolulu. Honniball i Lucey analitzaren les dades obtinguts. La reducció de les dades a unitats físiques de flux fou realitzada per Sachin Shenoy, del Space Science Institute, de Boulder. Shuai Li, del Hawaii Institute of Geophysics and Planetology realitzà el calibratge entre l’absorció i l’abundància d’aigua i les dades M3. Charles A. Hibbits, de l’Applied Physics Lab, de la Johns Hopkins University, de Laurel, aportà dades de laboratori sobre la banda de 6 μm en meteorits. Thomas M. Orlando, de la School of Chemistry and Biochemistry del Georgia Institute of Technology, d’Atlanta, i William M. Farrell, del NASA Goddard Space Flight Center, de Greenbelt, assessoraren Honniball i Lucey en les energies d’activació, coberta de monocapa i estimacions de superfície. Dana M. Hurley, de l’Applied Physics Lab de Laurel participà en la interpretació dels resultats pel que fa a l’exosfera lunar.
Les observacions es feren el 31 d’agost del 2018 en el NASA/DLR SOFIA, projecte operat conjuntament per la Universities Space Research Association, Inc. (USRA) com a contractista de la NASA i pel Deutsches SOFIA Institut com a contractives de la DLR. L’instrument utilitzat fou el Faint Object infraRed CAmera (FORCAST), que té un rang de longituds d’ona de 5 a 8 μm i una resolució espectral de 200, així com una elevada ratio de senyal:soroll. L’escletxa d’entrada que defineix la porció de Lluna observada és de 2,4 x 191 segons d’arc, corresponent a 248 píxels. L’extensió espacial en el centre del disc lunar és de 4,8x1,5 km2, per bé que la resolució especial en les vores del disc és inferior.
El 31 d’agost del 2018 la lluna era minvant, amb un angle de fase de 57,5°. Les regions observades en aquella sessió foren:
- una zona de latitud alta de l’hemisferi sud prop del cràter Clavius.
- una zona de latitud baixa dins de Mare Serenitatis.
Com era d’esperar, el senyal d’emissió de 6 μm de la regió del Clavius és prou forta quan se la compara amb la de Mare Serenitatis. Tots els espectres de la regió del Clavius mostren aquesta emissió, i la majoria (98%) depassen en 2 sigmes el soroll de fons, per bé que tan sols un 20% arriben a les 4 sigmes.
A través de Hibbits, s’estudiaren en el laboratori altres materials planetaris pel que fa a la banda de 6 μm. Honniball et al. també compararen les mesures obtingudes amb la bibliografia sobre hidrats cristal·lins, vidre del basalt de la dorsal oceànica i meteorits. Les bandes de tots aquests materials coincideixen prou com per assumir que el tret es correspon a la presència d’aigua molecular.
L’abundància d’aigua
Li, a través d’una anàlisi de laboratori de seccions fines de material geològic, obtingué la relació entre l’altura del pic d’absorbància de 6 μm i l’abundància de l’aigua. Gràcies a aquesta relació, Honniball et al. estimen que les abundància d’aigua en la regió de Clavius van de 100 a 400 ppm, amb una mitjana de 200 ppm.
La comparació amb les dades del Moon Mineralogy Mapper (M3) suggeriria que l’H2O seria la principal espècie hidroxílica.
D’on provindria aquesta aigua? Honniball et al. enumeren: 1) la quimioabsorció sobre superfícies de grans d’aigua present en l’exosfera lunar; 2) la introducció a través de micrometeorits rics en materials volàtils; 3) reaccions químics d’hidroxils que generarien aigua in situ sobre la superfície de grans; 4) la formació d’aigua in situ a partir d’hidroxils aportats per micrometeorits. De tota manera, Honniball et al. consideren que l’aigua detectada s’hauria de trobar dins de materials vitris de la superfície. El 30% del sòl lunar consisteix en vidres derivats de l’impacte de meteorits, de manera que hom esperaria en aquest material vitri l’abundància d’aigua fos de mitjana de 700 ppm.
Per tot plegat, caldria pensar que tot aquesta aigua es genera a través d’un mecanisme associat amb impactes de micrometeorits, que transformaria hidroxils lunars en aigua segrestada dins de vidres. Aquest procés seria més eficient en latituds altes que no pas en les regions equatorials.
De moment, tan sols s’ha rastrejat la regió del cràter Clavius. Observacions d’altres regions, i realitzades en diferents moments de la lunació, ajudarien a fer-se un quadre més precís de tota aquesta aigua molecular.
Lligams:
- Molecular water detected on the sunlit Moon by SOFIA. C. I. Honniball, P. G. Lucey, S. Li, S. Shenoy, T. M. Orlando, C. A. Hibbitts, D. M. Hurley, W. M. Farrell. Nature Astronomy (2020).
- NASA’s SOFIA Discovers Water on Sunlit Surface of Moon, comunicat de premsa de la NASA.