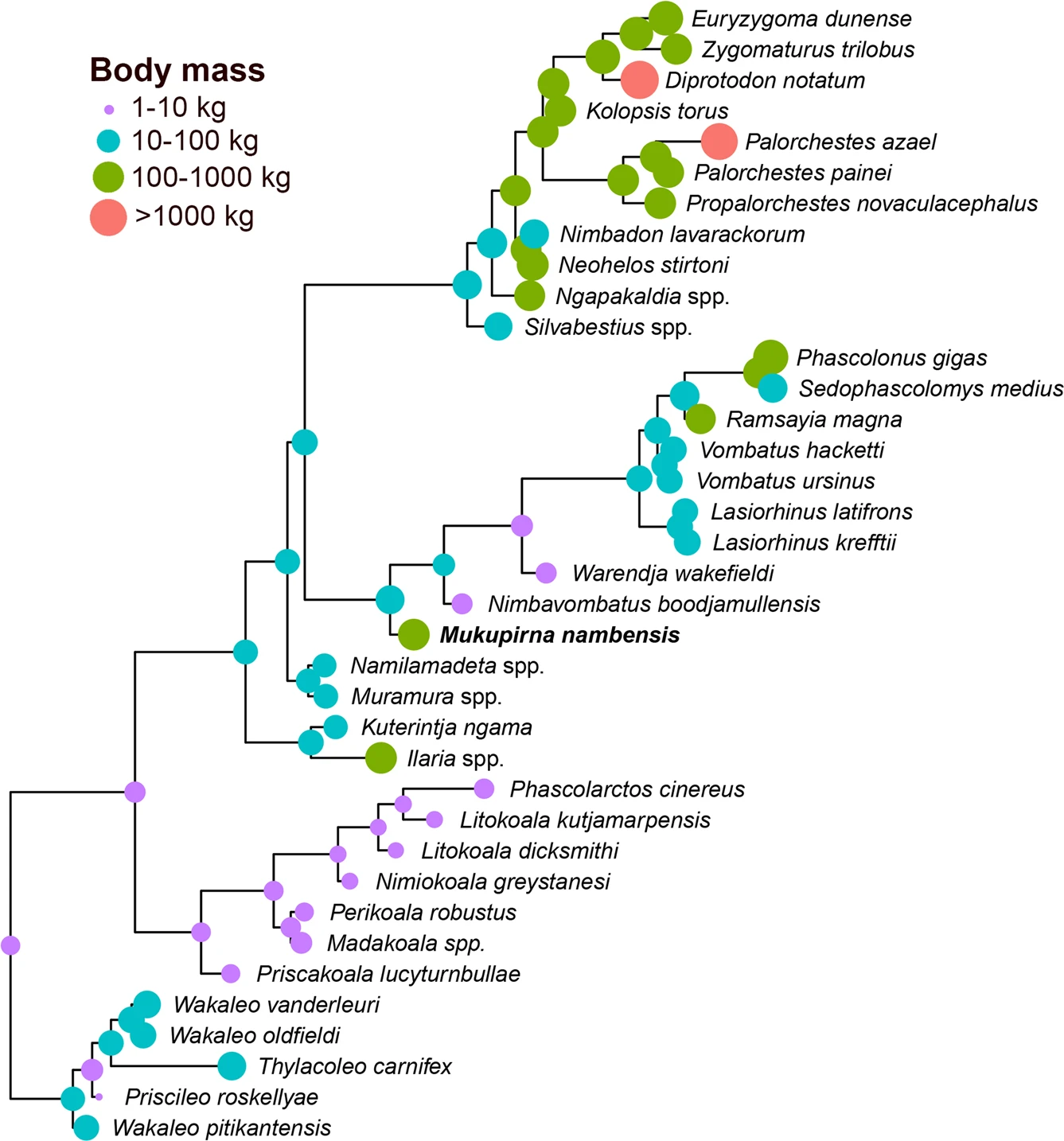
Fotografia de Brian W. Schallet que mostra un dofí mular indopacífic al Shark Bay Marine Park
Els dofins de la Badia dels Taurons
Aquest estudi fou concebut per Michael Krützen (del Departament d’Antropologia de la Universitat de Zürich), Simon J. Allen (de la School of Biological Sciences de la University of Bristol) i Sonja Wild (investigadora predoctoral de la Faculty of Biological Sciences de la University of Leeds). Les anàlisis estatístiques foren realitzades per Wild i William J. E. Hoppitt (de la University of Leeds). Wild recollí dades i redactà el primer esborrany.
La recerca fou finançada a través dels projectes que Krützen té amb la Fundació Nacional Suïssa per la Ciència, la SWRRFI d’Austràlia, la National Geographic Society, A.H. Schultz Stiftung, Claraz-Schenkung, Julius-Klaus Stiftung, i W.V. Scott Charitable Trust.
Els autors tenen paraules d’agraïment per a la Shark Bay Salt Pty. Ltd., i pels estudiants i voluntaris que contribuïren a la recollida de dades per a la Dolphin Innovation Project i la Shark Bay Dolphin Research.
Els subjectes d’aquest estudi són els dofins mulars indopacífics (Tursiops aduncus) que viuen de manera salvatge en el golf occidental de la Shark Bay, a Western Australia. Les dades genètiques s’obtingueren a través d’un sistema remot de presa de biòpsies, dins del marc del permisos concedits pel Department of Biodiversity, Conservation and Attractions i el Department of Agriculture and Food. Els protocols foren aprovats pels comitès d’ètica animal de la University of Western Australia, la Murdoch University i la Universitat de Zuric.
Les dades utilitzades foren recollides entre el 2007 i el 2018, dins de les observacions fetes des de vaixell, amb la base en la població de Useless Loop. Cada vegada que el vaixell s’acostava a un grup de dofins, els observadors prenien les dades de localització (per GPS) i determinaven la composició del grup a través de la identificació fotogràfica d’individus. Els individus que es trobaven a menys de 10 metres d’un altre dofí durant els primers cinc minuts de cada encontre eren considerats part del mateix grup. Tots els casos de comportament de “shelling” (la captura d’una closca de gasteròpode marí per consumir els peixos que hi haguessin amagats a dins) eren anotats, indicant la identitat del dofí que ho feia. Aquests individus rebien la designació de “shellers”.
Les biòpsies es prenien de forma oportunista després de cinc minuts d’observacions comportamentuals. S’adreçaven a individus dels que no s’hagués tingut mostra prèvia. Per prendre-la utilitzaven un rifle modificat del calibre 0,22, amb el que disparaven dards de policarbonat amb puntes d’acer inoxidable. Els teixits recollits d’aquesta manera eren guardats en una solució de dimetil-sulfòxid (20%) saturada amb clorur de sodi, a una temperatura de 4°C. En arribar al laboratori, aquestes mostres eren dutes al congelador de -80°C.
En total s’extragué ADN de 295 mostres. El genotipat es feia per tres PCR múltiplex d’un total de 27 loci microsatèl·lits, acoblada amb una seqüenciació ulterior.
Del 293 mostres s’obtingué un genotip per a 24-27 loci, i fou a partir d’elles que es calculà el grau de relació biparental diàdica. Així es podia identificar si dos individus eren mig germans (relació de 0,25), pare i fill (relació de 0,5), germans complets (relació de 0,5), cosins germans (relació de 0,125) o si no tenien cap parentiu (relació de 0). Com que els hàbits de depredació en els dofins mulars indopacífics s’aprenen bàsicament de la mare, de les mostres d’ADN es realitzà una amplificació del fragment D-Loop de l’ADN mitocondrial (d’herència materna) per tal d’assignar cada individu a un llinatge matern. En les mostres analitzades s’identificaren els haplotips A, B, D, E, H, F, I i K. A nivell d’anàlisi, aquests haplotips es condensaren en tres haplogrups: E, D i “altres”.
Al mateix temps, de cada mostra individual es feien PCR dels loci ZFX i SRY per tal de determinar el sexe de l’individu.
En l’anàlisi de difusió basat en xarxa (NBDA) cal combinar una bona mida mostral amb la qualitat de dades. D’aquesta manera, convé excloure individus dels quals tan sols s’han fet unes poques observacions, per tal que la qualitat de les dades augmenti encara que sigui en detriment de la mida mostral. Així, Wild et al. tan sols tingueren en compte les de 538 individus, dels quals s’havien fet un mínim de cinc avistaments.

Localitzacions de les observacions utilitzades en aquest estudi
L’hàbit de pescar amb closques buides
Entre el 2007 i el 2018, en el marc de 5278 encontres amb grups de dofins, s’identificaren un total de 1.035 individus diferents. En aquests observacions es detectaren 42 casos de consum de closca (“shelling event”), realitzats per 19 individus de tres llinatges mitocondrials diferents. Certament, el “shelling” és un hàbit rar, però Wild et al. alerten que és un comportament que se sol despatxar en pocs segons i que, per tant, és fàcil que passi desapercebut als observadors. Per això tingueren en compte una submostra de 310 individus pels quals hi havia hagut un mínim d’11 observacions. D’aquests 310 individus, 15 eren “shellers” (6 femelles i 9 mascles).
Per investigar la via de transmissió d’aquest comportament, Wild et al. realitzaren una anàlisi de difusió d’ordre d’adquisició. En aquesta tècnica es combinen les dades de xarxa social (el fet que dos individus pertanyin al mateix grup), d’entorn (el fet que dos individus es trobin en ambients semblants) i de genètica (el fet que dos individus tinguin una relació de parentiu). D’acord amb aquests models, els resultats indicarien que la transmissió d’aquest comportament seria eminentment social. Que el comportament sigui “descobert” de manera individual, o que sigui “heretat”, no encaixa amb les dades. Tampoc no seria un aprenentatge vertical, transmès per la mare a la progènie. Quedaria, doncs, una transmissió social no-vertical, incloses la transmissió horitzontal (entre individus de la mateixa generació) o l’obliqua (entre un individu d’una generació anterior i un de més jove).
La transmissió social del “shelling” semblaria més efectiva en grups petits que no pas en grups més grans. Aquesta transmissió social sembla independent de factors com el sexe, l’haplotip mitocondrial o la fondària marina. Existeixen, però, diferències en la velocitat d’aprenentatge relacionada amb l’haplotip, de manera que els dofins amb haplotip D i E ho fan molt més ràpidament que els altres haplotips.
Wild et al. ofereixen la primera prova quantitativa d’un comportament alimentari de transmissió no-vertical en un odontocet. Caldrà ampliar la base de dades d’observacions per poder distingir si és una transmissió obliqua o horitzontal. Pel que se sap, aquesta transmissió dependria de la durada i freqüència de proximitat entre el “demostrador” i l’“aprenent”, i seria més efectiva en grups més petits, on les relacions interindividuals serien més focalitzades. En els dofins de Shark Bay, aquestes relacions més estretes són típiques entre mascles de grups socials més cohesius, però les femelles també mostren alts nivells de tolerància social.
Wild et al. contrasten la pràctica del “shelling” amb la de l’“sponging”. L’“sponging” s’aprèn de manera vertical, sobretot apresa per les filles a partir de la mare, en el marc de l’adopció de les mateixes àrees de pesca (els dofins mulars de Shark Bay són en aquest sentit matrilocals). El “shelling”, però, seria una estratègia més oportunista, que no requereix l’especialització de l’“sponging”.
La recopilació de dades esclarirà aspectes com ara el percentatge de dofins que adquireixen el “shelling” de manera asocial, és a dir de forma individual (Wild et al. ho estimen en un 43%). El “shelling”, com el “sponging”, són exemples d’ús d’eines de pesca per part dels dofins. En el “shelling” el dofí utilitza la closca buida d’un gasteròpode marí per atrapar preses. Després de mantindre-la submergida, el dofí treu la closca de l’aigua, i la sacseja per fer caure el peix o peixos que hi hagin entrat.
Lligams:
- Integrating Genetic, Environmental, and Social Networks to Reveal Transmission Pathways of a Dolphin Foraging Innovation. Sonja Wild, William J.E. Hoppitt, Simon J. Allen, Michael Krützen. Current Biology (2020).
- Shark Bay Dolphin Research.