Arqueologia: Heiko Prümers, del Deutsches Archäologisches Institut, és el primer autor d’una recerca feta amb la tecnologia òptica de teledetecció LIDAR que posa al descobert l’urbanisme de baixa densitat de l’Amazònia Boliviana vinculat a la cultura de Casarabe. En l’article aparegut avui a Nature remarquen la presència de dues localitats grans (de 147 i de 315 hectàrees) en un sistema d’assentament quàdruple. El Lidar ofereix la possibilitat d’investigar les restes arqueològiques d’urbanisme agrari de baixa densitat ara amagat sota els boscos tropicals: l’Amazònia, doncs, presentava un desenvolupament que ha estat detectat al sud-est d’Àsia, a Sri Lanka o a Mesoamèrica. L’àrea de la cultura Casarabe (desenvolupada entre l’any 500 d.C. i 1400 d.C.) ocuparia, segons les dades arqueològiques actuals, una superfície de 4500 km2. Un dels assentaments descrit per Prümers et al. controlaria un àrea de 500 km2. Els dos assentaments presenten una arquitectura civil i cerimonial amb plataformes inclinades al capdamunt d’estructures en forma d’U, monticles acabats en plataformes rectangulars i piràmides còniques de fins a 22 metres d’alçada. Aquests assentaments són encerclats per bancs poligonals concèntrics, i connecten amb camins rectes i elevats amb altres assentaments menors situats a diversos quilòmetres. Canals i embassaments integraven una infrastructura hidrològica al servei d’aquests assentaments. Tot plegat, doncs, suposava un paisatge modificat antropogènicament en allò que Prümers et al. qualifiquen d’urbanisme tropical de baixa densitat.
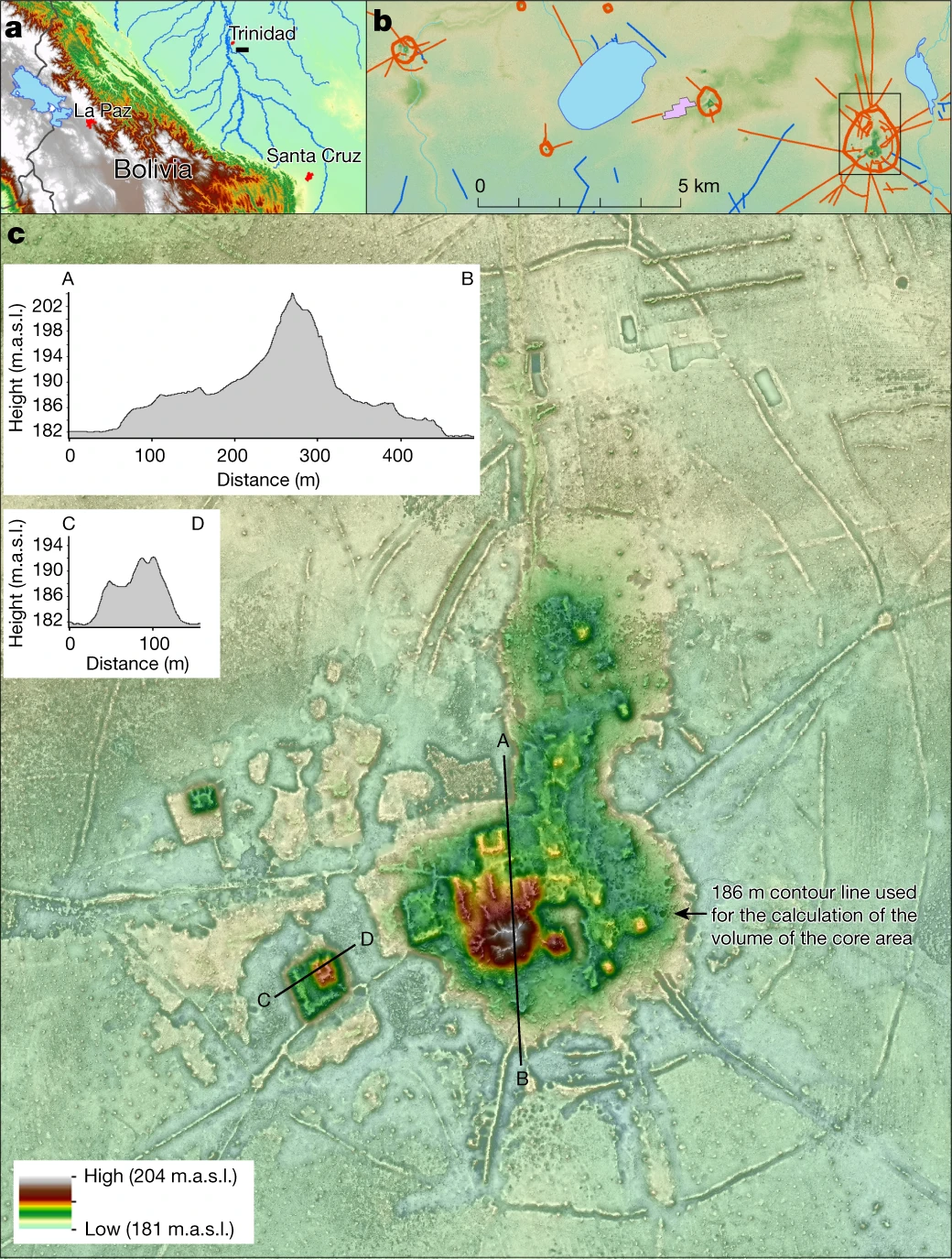
L’assentament de Cotoca
Urbanisme tropical de baixa densitat
Aquesta recerca fou dissenyada per Heiko Prümers (Deutsches Archäologisches Institut, Kommission für Archäologie Aussereuropäischer Kulturen, Bonn), Carla Jaimes Betancourt (del Departament d’Antropologia de les Amèriques de la Universitat de Bonn) i José Iriarte (Department of Archaeology, College of Humanities, University of Exeter). El treball de camp fou conduït per Prümers, Jaimes, Iriarte i Mark Robinson (Exeter). Les dades crues de lidar foren processades per Martin Schaich (de ArcTron 3D GmbH). Prümers realitza la feina cartogràfica, estatística i de càlcul. En l’anàlisi de dades participaren Prümers, Jaimes, Iriarte i Robinson. Prümers, Jaimes, Iriarte i Robinson redactaren l’article, que fou tramès a Nature el 10 de juliol del 2021. Amb Prümers com a autor corresponsal s’inicià un procés de revisió, amb Chris Fisher, Roland Fletcher, John Walker i un altre revisor. Aquest procés de revisió conclogué amb acceptació el 20 d’abril, i l’article fou publicat el 25 de maig.
Els autors tenen paraules d’agraïment pel suport del Govern Autònom Departament de Beni i del Ministeri de Cultura i Turisme de Bolívia. Agraeixen també la feina a Skyplus (E. Méndez P. i G. Nogales H.) de Santa Cruz de la Sierra, i l’aportació de R. Landivar sobre la toponímia actual. El treball es finançà amb aportacions de l’Institut Arqueològic Alemany i el servei de Intervenciones Urbanas del Ministeri de Planificació de Bolívia. Les dades de lidar es recolliren amb el sensor VUX-1 utilitzat en el projecte ERC-PAST d’Iriarte.
Les poblacions agrícoles de Llanos de Mojos, durant l’Holocè Tardà, transformaren 120.000 km2 del sabana amazònica d’inundació estacional en un paisatge productiu. L’establiment de sistemes de control d’aigua fou acompanyat amb una nova organització sociopolítica. Al sud-est d’aquesta regió, es beneficià de la qualitat del sòl i d’una topografia més elevada. És en aquesta zona de 4500 km2 on es desenvolupà, entre els segles VI i XIII, l’anomenada cultura Casarabe.
Campanyes amb sensors remots i anàlisis de reconeixement de camp havien revelat l’existència de 189 grans assentaments monumentals (les ‘lomas’), 237 assentaments menors i 957 km de canals i carreteres. Les lomas eren habitades tot l’any per poblacions agrícoles dedicades al cultiu de panís (Zea mays) i altres collites, complementat amb activitat caçadora i pesquera.
Prümers et al. conduïren una cartografia aèria per làser de sis àrees (d’un total de 204 km2) corresponents a les zones de més densitat de lomas. La tècnica Lidar (“light detection and ranging”) els permet documentar en detall dos grans assentaments i vint-i-quatre de més petits. De fet, dels 24 assentaments menors, tan sols 9 havien estat descrits prèviament.
Prümers et al. aventuren una classificació jeràrquica dels assentaments en quatre nivells atenent a les dimensions de les plataformes, el grau d’elaboració de l’arquitectura civil i cerimonial, l’abast de la perifèria poligonal, el nombre de carreteres que hi condueixen, l’escala de la infrastructura hidrològica.
La metodologia lidar
La tecnologia lidar permet accedir a jaciments arqueològics actualment amagats per denses boscúries. En el 2011 fou emprada en la prospecció arqueològica d’Iténez/Guaporé en el marc del Projecte Germano-Bolivià de Mojos (PABAM), i després ho ha estat en zones amazòniques de Brasil i Perú.
En aquest treball s’emprà un escàner Riegl VUX-1, amb un Trimble APX-15 UAV GNSS, instal·lar en un Eurocopter AS350. La taxa de repetició de pols era de 200 kHz. L’alçada de vol era de 200 metres, i la velocitat de 45 nusos. Els vols es feien en franges paral·leles de 200 metres, amb un 50% de superposició. El post-processament de dades es feia amb el programari RiAnalyze d’ArcTron.
Els volums de terra mobilitzada en la construcció d’estructures fou estimat a El Cerrito, a la Loma Landívar i a la Loma Cotoca amb ArcGIS.
Hom disposa de 144 datacions per radiocarboni de la cultura Casarabe, centrada en la informació estratigràfica de la Loma Alta de Casarabe, Mendoza, Salvatierra i Pancho Román. Encara que la cronologia més acceptada de la cultura Casarabe va de l’any 500 d.C. al 1400 d.C., hi ha dues datacions anteriors (segles I-II d.C.).
Els grans assentaments
Són assentaments de mides superiors a les 100 hectàrees. Els dos més importants són Cotoca (de 147 hectàrees) i Landívar (de 315 hectàrees), l’abast dels quals tan sols ha estat posat de manifest per les dades lídar de Prümers et al.
Aquests dos assentaments són encerclats per tres estructures defensives concèntriques, consistents en un fossat i un baluard, en alguns casos de doble muralla. A Cotoca tan sols es conserva l’estructura defensiva interior en alguns sectors, cosa que Prümers et al. atribueixen a una adaptació al creixement de l’assentament.
Edificacions de terra (plataformes, algunes en forma d’U; piràmides còniques) arriben a alçar-se més de 20 metres per damunt de la sabana. La base d’aquestes edificacions són terrasses artificials de fins a 6 metres d’alçada. Tant a Cotoca com a Landívar, aquestes edificacions s’orienten en sentit nord-nord-oest, tal com ho fan també els enterraments estesos de la cultura Casarabe.
El nucli de Cotoca, definit per aquesta terrassa artificial, té una extensió de hectàrees, més del triple de la dels centres secundaris. La terra de les construccions fou adquirida, si més no en part, d’àrees adjacents al centre de l’assentament. Així a Cotoca s’excavà una franja de 50 a 80 metres d’ample que envoltava la terrassa central. En l’actualitat, durant l’estació de pluges, la franja s’omple d’aigua, i roman de fet pantanosa durant bona part de l’estació eixuta.
Cotoca i Landívar eren els centres primaris d’una xarxa regional d’assentaments connectada per carreteres rectes encara visibles en l’actualitat. En indrets estratègies de les carreteres hi ha plataformes de 20 x 25 metres i 2 metres d’alçada que, segons Prümers et al., serien llocs de vigilància, de forma que l’accés als grans assentaments era restringit i controlat.
La mobilització de terres en la construcció de Landívar és estimada en 276.000 m3. En el cas de Cotoca la xifra arribaria a 570.690 m3. A tall de comparació, Prümers et al. recorden que en la construcció d’Akapana, l’estructura més gran de Tiwanaku, es mobilitzaren 53.546 m3 de terra.
Centres secundaris
Centres secundaris com El Cerrito i Salvatierra es caracteritzen per plataformes de base amb superfície de 2 a 6 hectàrees. Un tancament poligonal circumscriu al voltant un àrea de 21 a 41 hectàrees. L’arquitectura civil i cerimonial consisteix en un o diversos monticles acabats en plataforma.
Els centres terciaris consisteixen en una plataforma base de 0,5 hectàres, amb una sol monticle, i una rasa circular que abasta una àrea màxima de 2,5 hectàries.
Els centres de quart nivell consisteixen en petits llocs elevats (0,34 hectàrees de mitjana). Aquestes illes en mig del bosc eren probablement utilitzades com a campaments temporals o es dedicaven a activitats especialitzades.
Prümers et al. no descarten l’existència encara d’un cinquè nivell de petits llogarrets. Ara bé, com aquests llogarrets no tindrien associats arquitectura de monticle, no són detectables per lidar.
Prümers et al. remarquen el caràcter integrat, continu i dens dels assentaments de la cultura Casarabe. En la zona de 4500 km2 hi ha una mitjana de 10 assentaments primaris, secundaris o terciaris en un radi de 10 km al voltant de cada assentament. La densitat augmenta en el sector oriental, on la mitjana de distància entre assentaments cau a 2-4 km. Carreteres i canals connecten assentaments formant clústers de 100 a 500 km2. La xarxa és força centralitzada, de manera que els assentaments terciaris no tenen connexions directes entre ells, sinó que passen a través dels assentaments secundaris i primaris.
Cotoca és el centre d’un àrea de 500 km2, dividida gairebé a la meitat entre bosc i sabana, que inclou 18 assentaments monumentals (3 centres secundaris, 2 terciaris i diversos clústers d’assentaments quaternaris en direcció sud-est i oest). Els canals de Cotoca connecten amb el riu Ibare al sud i amb llacs a l’est. Un canal de 7 km de longitud aportava aigua de la Laguna San José.
No sempre l’assentament principal es troba al mig de la regió circundant, ni tot el territori era dominat per assentaments principals. L’assentament secundari de El Cerrito dominava un àrea de 100 km2.
La cultura Casarabe en el context de l’Amazònia occidental
Prümers et al. consideren que les dades aportades descarten la visió d’una Amazònia pre-colombina de poblament espars. La cultura Casarabe modificà el paisatge social i públic a través de la monumentalitat. Seria el primer cas conegut d’urbanisme agrari tropical de baixa densitat d’Amèrica del Sud. En termes d’abast i monumentalitat no queden enrere de les cultures andines. Val a dir que al sud d’Amazònia també hi havia assentaments sofisticats i interconnectats, però allà no es detecta l’arquitectura monumental de la cultura Casarabe.
Lligams:
- Lidar reveals pre-Hispanic low-density urbanism in the Bolivian Amazon. Heiko Prümers, Carla Jaimes Betancourt, José Iriarte, Mark Robinson, Martin Schaich. Nature (2022)