Filogenòmica: El Regne Animal de la classificació tradicional fou dividit per Ernest Haeckel el 1874 entre “protozous” i “metazous”, és a dir bàsicament entre “animals unicel·lulars” i “animals pluricel·lulars”. En les dècades següents les esponges foren considerades com un exemple de “parazous”, és a dir situats en el mig de les dues categories. Al capdavall, les esponges (o porífers) presenten un pla corporal relativament senzill. D’aquesta manera, hom pensava que serien el “grup germà” de la resta de “metazous”, és a dir dels “eumetazous”. Amb la inclusió dels “protozous” en el Regne dels Protoctistes, la terminologia se simplifica en el sentit de redefinir els “Animalia” com a sinònim de “Metazoa”. Però la qüestió de les relacions entre els llinatges principals d’Animalia no és gens resolta. Les classificacions del segle XX feren proliferar una mà de “phyla” o “divisions” de les quals els “Porifera” (les esponges) en serien la primera. Aquestes classificacions reflectien precisament la dificultat d’entendre l’origen i l’evolució de trets animals: pluricel·lularitat, simetria bilateral, etc. L’adveniment de la biologia molecular a partir dels anys 1960 semblava obrir noves perspectives que s’unirien a les evidències aportades per l’anatomia comparada, l’embriologia o la paleontologia. Ara bé, estudis genòmics indicaven de vegades resultats paradoxals, com ara que els ctenòfors (meduses, pòlips, coralls) eren realment el grup germà extern a la resta d’animals, mentre que les esponges hi serien més acostades. Això implicava que les esponges actuals serien una simplificació de la forma ancestral. Anthony K. Redmond i Aoife McLysaght del Molecular Evolution Lab del Smurfit Institute of Genetics de la Universitat de Dublín, mostren en un article a Nature Communications com la incorporació de models de barreja heterogènia de lloc i de filogenòmica particionada indicaria veritablement que les esponges són el grup germà de la resta d’animals pluricel·lulars. Amb aquesta aproximació redueixen errors sistemàtics dels models filogenètics més habituals.
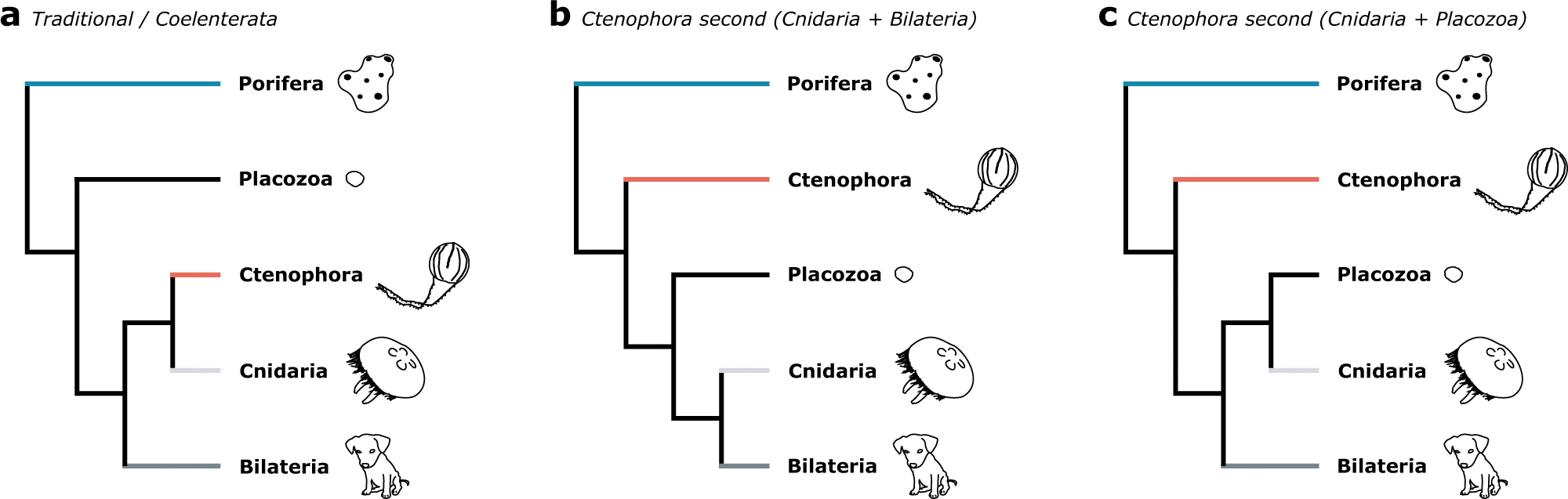
Si constatem que els porífers són el grup germà de tota la resta de metazous, queden obertes les qüestions sobre com es relacionen entre ells placozous, ctenòfors, cnidaris i bilateris. El laboratori de McLysaght es proposa ara resoldre quins d’aquests tres esquemes és el més plausible.
El lloc dels porífers en la classificació animal
Anthony K. Redmond concebé i dissenyà aquest projecte de recerca, i conduí totes les anàlisis. Juntament amb Aoife McLysaght, la cap del Laboratori d’Evolució Molecular de l’Institut Smurfit de Genètica, analitzà les dades i redactà l’article. Els dos autors tenen paraules d’agraïment per a Hervé Philippe i Nicolas Lartillot per compartir les dades emprades dels projectes BEA, LEAN i LEAP, i per a Daniel J. Macqueen per l’orientació inicial de l’estudi. L’estudi fou finançat amb el projecte 771419 de l’European Research Council.
L’article fou tramès a Nature Communications el 7 d’agost del 2020. Acceptat el 24 de febrer, fou publicat el 19 de març.
Les esponges van rebre el 1836 de Robert Edmond Grant (1793-1874) la designació de “Porifera” pels porus i canals que permeten la circulació d’aigua a través dels seus cossos. En les dècades ulteriors, les esponges foren considerades ja com els animals pluricel·lulars més simples. D’una banda, manquen de teixits veritablement dits, d’òrgans, i de sistemes diferenciats (sistema nerviós, digestiu o circulatori). De l’altra, no presenten simetria corporal. D’ací que en l’esquema de “protozous” i “metazous”, animals primitius i animals evolucionats, se’ls reservés la categoria de “parazous”, com a animals d’una pluricel·lularitat incipient.

En aquesta imatge de les Illes Caiman podem veure quatre espècies d’esponges: ‘Aplysina fistularis’ (tubular groga), ‘Niphates digitalis’ (vasal porpra), ‘Spiratrella coccinea’ (incrustant roja) i ‘Callyspongia’ (corda grisa).
Aquesta concepció heretava la idea de l’escala de la natura. El graó següent als porífers seria ocupat pels ctenòfors, un fílum definit per Eschscholtz el 1829, que sí presenta simetria rotacional, a més d’una diferenciació en teixits: amb musculatura, budell i sistema nerviós.
Fotografia de Marsh Youngbluth que ens mostra un exemple de ctenòfor mesopelàgic, ‘Bathocyroe fosteri’
L’adveniment de tècniques de biologia molecular fou aplicat a resoldre amb més detall l’esquema de classificació dels diferents phyla animals, amb l’esperança de reduir-ne el nombre. Certament, ens han ajudat força, però en el cas de la relació entre porífers, ctenòfors i la resta d’animals, han vingut a somoure conceptes ja establerts. En particular, cal resoldre quin dels dos grups (o si ho és un tercer) té una relació externa (de grup germà) amb el gros de metazous. Les dades clàssiques assenyalaven les esponges, però estudis genètics fan pensar que el lloc realment l’ocupen els ctenòfors.
La qüestió té rellevància sobre l’origen del sistema nerviós, de la musculatura, i d’altres trets fonamentals (simetria bilateral, etc.). Si les esponges no són el grup germà, hom pot pensar que són el resultat d’una simplificació: que en el curs de la seva evolució han perdut el sistema nerviós, la musculatura i la simetria. O, alternativament, que aquestes característiques han aparegut de manera independent en un mínim de dues ocasions: per als ctenòfors i per als altres metazous.
En aquestes dècades de recerca ha augmentat també el nostre coneixement d’altres grups de “parazous” com ara els placozous. Però alhora això complica el debat sobre la seva posició en aquest esquema.

‘Trichoplax adhaerens’, un exemple de placozou, de 0,5 mm de mida.
Redmond & McLysaght recorden que hi ha factors que poden conduir a estimacions errònies en els estudis filogenètics, com ara la no-independència de caràcters, l’heterogeneïtat en el procés de substitució genètica (heterotàquia, heteropecíl·lia). Per això mateix, en els darrers anys, s’han desenvolupat models de barreja infinita que tinguin en compte aquesta heterogeneïtat, com el model CAT o el model CATGTR. Per exemple, aquests models poden detectar millor una saturació de substitucions, en les que s’amagarien substitucions múltiples que poden conduir a una evolució convergent de posicions en les seqüències nucleica o aminoacídica. En la comparació de seqüències aminoacídiques també és rellevant tractar els 20 aminoàcids possibles en els seus grups bioquímics. Una altra aproximació que refina les dades és una partició de les dades en grups de gens, per a cadascuna de les quals s’aplica un model separat.
L’aproximació metodològica de Redmond i McLysaght
Redmond & McLysaght fan ús de les dades utilitzades per Nicolas Lartillot: en LEAP i en LEAN s’incloïen 35373 llocs corresponents a 146 gens i 32 tàxons. En les dades de Henner Brinkmann (BEA) es tenien en compte 24.294 llocs, corresponents a 133 gens i 40 tàxons. També utilitzen les dades de Ryan et al. (REA) sobre 88.384 llocs (406 gens i 60 tàxons), i les de Whelan et al. (WEA15) de 23.680 llocs (89 gens i 62 tàxons). Exclogueren Xenoturbella bocki per la seva posició incerta, i tan sols inclogueren com a grups externs als metazous els coanoflagel·lats.
Utilitzaren IQ-tree (v.1.5.4-omp) per realitzar les comprovacions de model i les anàlisis filogenòmiques. Els models eren escollits d’acord amb el criteri d’informació bayesià.
Els models més adients
Redmond & McLysaght separen els models contemplats en quatre grups:
- T1: models homogenis de lloc estàndards, com JTT, WAG o LG.
- T2: models de barreja multi-matriu, com UL2, UL3, EX2, EX3, EHO, EX_EHO, LG4M.
- T3: models de barreja infinita com CAT.
- T4: model CATGTR.
En el conjunt de dades BEA, el grup extern són els arqueons. Les anàlisis estàndard de BEA condueixen a identificar Encephalitozoon cuniculi, un microsporidi, com a grup germà de tots els eucariotes. Això es degut a una alta taxa evolutiva. L’aplicació d’estratègies com descartar gens d’evolució ràpida, millorar el mostreig de tàxons o aplicar un model heterogeni de lloc com CAT corregeixen aquesta identificació errònia.
En els conjunts LEAP i LEAN s’inclouen fonamentalment tàxons de Bilateria, prenent a Fungi com a grip extern. Mentre que en LEAP hi ha presència de platihelmints, en LEAN hi ha presència de nematodes. En aquests conjunts, l’aplicació de models homogenis de lloc condueix a la conclusió espúria que artròpodes i deuterostomis constitueixen un clade de celomats. Això es rectifica amb la inclusió de grups externs més propers, amb la millora del mostreig de grups interns i amb l’ús de models heterogenis de lloc: llavors sí apareix identificat com a monofilètic el grup dels protostomis (amb artròpodes i platihelmints) i el subgrup d’ecdisozous (amb artròpodes i nematodes).
Així doncs, els models heterogenis de lloc s’adapten millor a les particions de gens d’aquests conjunts de dades que no pas els models estàndards. La combinació de particions amb models homogenis és susceptible d’errors, però un excés de partició (en la que el gen és emprat com a unitat bàsica) també pot arrossegar a error els models heterogenis de lloc. De tota manera, la filogenòmica particionada combinada amb models heterogenis és suficient per indicar que el grup extern als “eumetazous” no serien els ctenòfors, sinó els porífers.
Pel que fa a altres relacions, les reanàlisis de Redmond & McLysaght desencoratgen contemplar els Xenacoelomorpha com a grup germà de tots els altres Bilateria. Pels que fa als Placozoa, aquestes re-anàlisis indicarien que són el grup germà dels Cnidaria (meduses, pòlips, coralls). Pel que fa a les relacions internes dels Porifera, les re-anàlisis indiquen que són un grup monofilètic.
El rol de la filogenòmica particionada en la resolució de l’arbre de la vida
Redmond & McLysaght consideren que la combinació d’una filogenòmica particionada amb la recodificació d’aminoàcids i models de barreja heterogènia té la capacitat de redreçar errors sistemàtics.
Però quin lloc ocupen realment els ctenòfors en l’arbre filogenètic dels animals? Tradicionalment, els ctenòfors havien estat considerat el grup germà dels cnidaris: així doncs els col·loblasts adhesius dels ctenòfors potser serien homòlegs als cnidoblasts irritants de meduses i pòlips. Justament, per això es creà el tàxon dels Coelenterata que incloïa ctenòfors i cnidaris. L’evidència morfològica i anàlisis de presència i absència de famílies de gens ha fet en els darrers temps recuperar aquest concepte de “celenterats”. La filogenòmica ho havia desencoratjat potser per una taxa evolutiva diferencial entre els ctenòfors (heteropecíl·lia). Si acceptem aquest esquema tradicional podem pensar que el sistema neural i la musculatura aparegué en l’ancestre comú de celenterats i bilateris.
Una altra alternativa és considerar els ctenòfors com el grup germà de tots els metazous no-esponges. Així, si les esponges són el “grup germà primer”, els ctenòfors serien el “grup germà segon” o més estret. Ara bé, en aquest escenari hauríem d’acceptar que trets com el sistema nerviós i la musculatura haurien aparegut en els ctenòfors de manera independent a cnidaris-bilateris. O bé, hauríem de pensar que els placozous, com a grup germà dels cnidaris-bilateris, haurien perdut aquestes característiques per evolucionar a una pluricel·lularitat més indiferenciada.
Per destriar aquestes possibilitats, Redmond & McLysaght consideren necessària una estratègia filogenòmica més sofisticada, que faci front a l’heterogeneïtat de les dades. A més, cal tindre present l’efecte que poden haver tingut les transferències gèniques horitzontals.
Lligams:
- Evidence for sponges as sister to all other animals from partitioned phylogenomics with mixture models and recoding. Anthony K. Redmond & Aoife McLysaght. Nature Communications 12: 1783 (2021).










