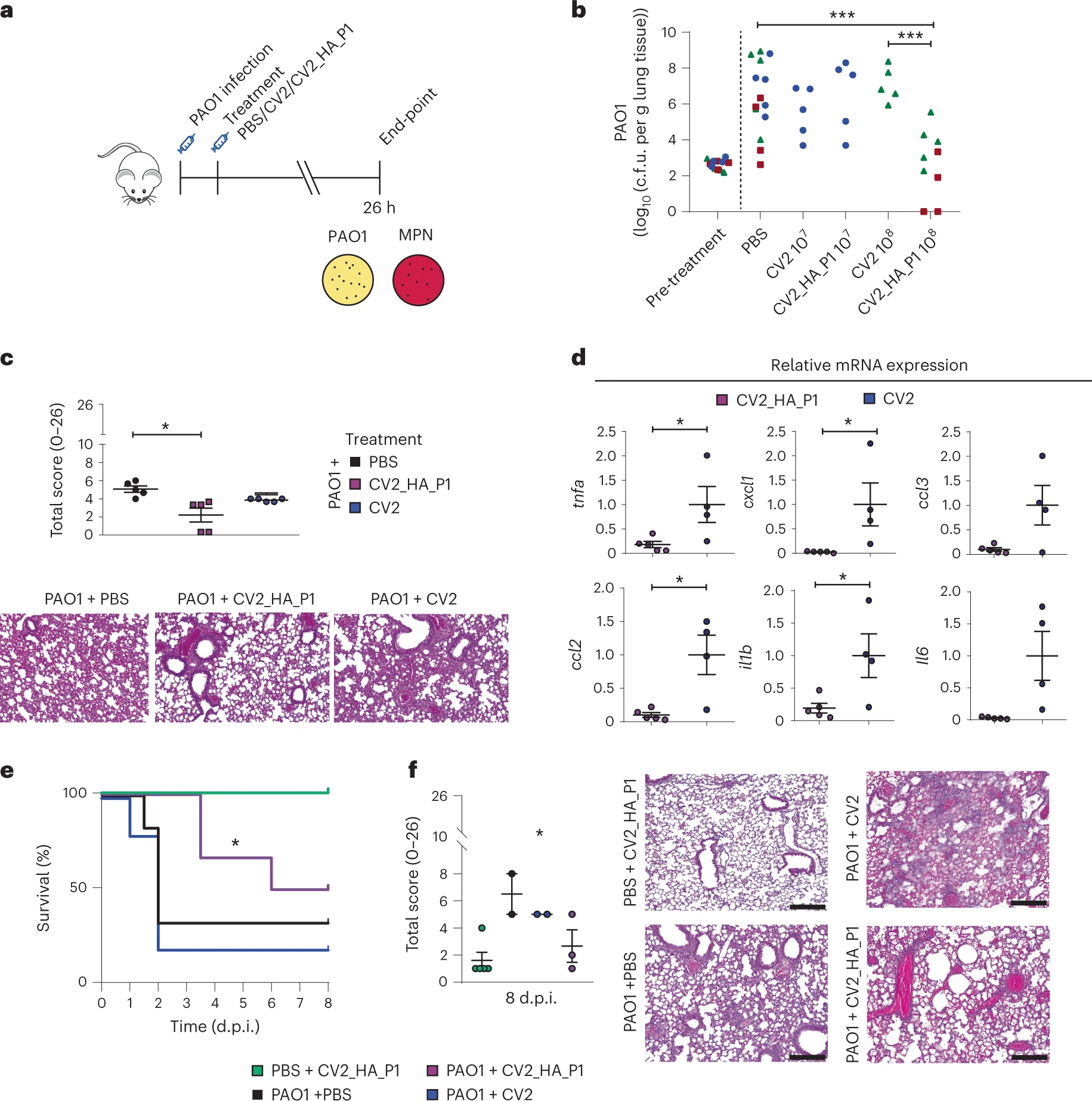
Genètica molecular: En llibres de text podem llegir, en l’enumeració de contrastos entre les cèl·lules eucariòtiques i les cèl·lules procariòtiques, que en les primeres les histones són els principals constituents proteics de la cromatina mentre que en les segones aquesta funció d’organització dels cromosomes la realitza d’una manera més laxa un seguit d’altres proteïnes. Val a dir però que les histones també tenen un rol important en la majoria d’arqueons, que són organismes tan procariòtics com els bacteris. Així que podem corregir la frase i dir que les histones són els constituents principals de cromatina en eucariotes i en la majoria d’arqueons. Però la cosa no acaba ací. L’anàlisi genòmica de les darreres dècades ha revelat que diversos genomes bacterianes codifiquen proteïnes amb possibles dominis de plegament d’histona. Molt s’ha discutit sobre si aquestes “proteïnes semblants a histones” són equivalents estructurals i funcionals a les veritables histones d’arqueons i d’eucariotes. En aquest debat irromp ara una publicació amb Antoine Hocher i Shawn P. Laursen com a primers autors en el que constaten que en els bacteris Bdellovibrio bacteriovorus i Leptospira interrogants les histones són components principals essencials de la cromatina. Indirectament dedueixen que les histones han de ser funcionalment rellevants en altres clades bacterians. L’anàlisi estructural del dímer la histona Bd0055 de B. bacteriovorus revela una topologia de plegament comparable a la descrita en arqueons i eucariotes. Ara bé, Bd0055 d’uneix a l’ADN d’una manera peculiar, formant-hi una beina de dímers estretament encaixats. Les histones dels genomes d’arqueons, eucariotes i virus, contràriament, són envoltades d’ADN. Hocher et al. conclouen que les histones són components integrals de la cromatina en els tres dominis de l’arbre de la vida, i que el domini dels bacteris no té res a envejar als altres en matèria d’innovació organitzativa.
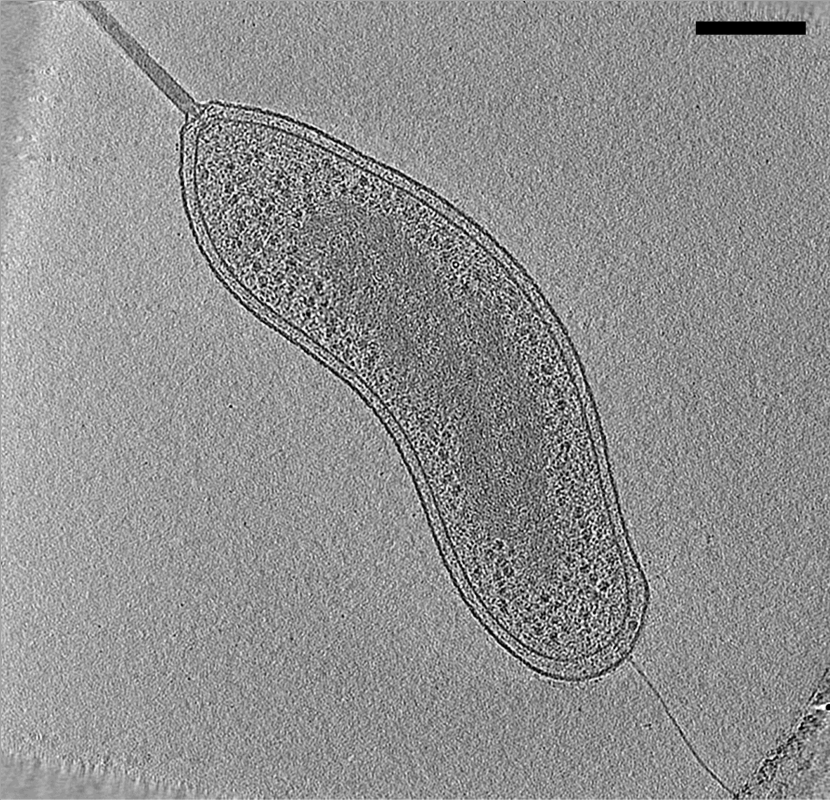
Electrofotografia d’una cèl·lula de “Bdellovibrio bacteriovorus”
Les histones
Els autors d’aquesta investigació són Antoine Hocher (del Medical Research Council London Institute of Medical Sciences i de l’Imperial College London), Shawn P. Laursen (del Department of Molecular, Cellular, and Developmental Biology de la University of Colorado, amb seu a Boulder), Paul Radford (de la School of Life Sciences de Nottingham), Jess Tyson (Nottingham), Carey Lambert (Nottingham), Kathryn M. Stevens (MRC i Imperial College), Mathieu Picardeau (Institut Pasteur), R. Elizabeth Sockett (Notthingham), Karolin Luger (de Colorado i del Howard Hughes Medical Institute) i Tobias Warnecke (MRC i Imperial College).
Hocher inicià aquest projecte i dugué a terme totes les anàlisis bioinformàtiques, els experiments de seqüenciació i de proteòmica, començant pels cultius bacterians i les anàlisi bioquímiques. Laursen realitzà tot el treball in vitro sobre la Bd0055 expressada en un recombinant d’Escherichia coli: caracterització biofísica, cristal·lografia, anàlisi estructural. Stevens ajudà en l’obtenció de dades d’homòlegs bacterians d’histones. Radford construí les soques de fusió de B. bacteriovorus Bd0055-mCherry i Bd0055-mCitrine, que foren fenotipades al microscopi per Tyson. Lambert i Radford intentaren les delecions del gen bd0055 en cultius de B. bacteriovorus tant depredadors com independents de presa. Picardeau aportà la biomassa per als estudis en l’espiroqueta L. interrogans de bioquímica i d’intent de deleció del gen LA_2458. Els projecte fou supervisat per Sockett, Luger i Warnecke. Hocher, Laursen, Luger i Warnecke dirigiren l’anàlisi i interpretació de dades, i redactaren l’esborrany que ara tenim a les mans.
Els autors agraeixen a la LMS Proteomics Facility la generació de dades d’espectrometria de masses, a la LMS Genomics Facility la seqüenciació d’ARN, i a Annette Ersbse (Colorado) l’assistència en la cristal·lografia. Radford, Tyson, Lambert i Sockett treballaren amb un fons de Wellcome. Hocher, Stevens i Warnecke amb un de UKRI MRC. Luger i Laursen són finançats pel Howard Hughes Medical Institute.
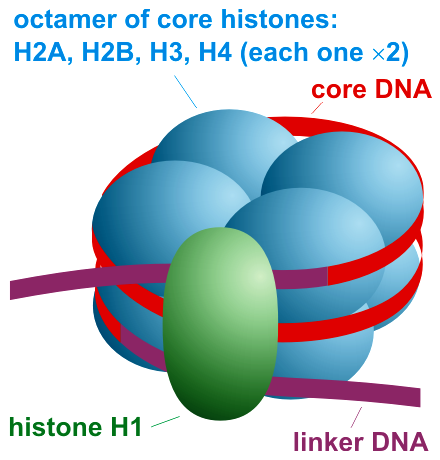
La unitat bàsica de la cromatina en eucariotes és el nucleasoma, consistent en un octàmer d’histones al voltant del qual s’aplega un segment de la doble hèlix d’ADN
La paraula “cromatina” designa el complex d’àcid desoxiribonucleic (ADN) i proteïna que constitueix el component principal del nucli de les cèl·lules eucariotes, és a dir de les cèl·lules que tenen un nucli envoltat per una doble membrana lipídica. La unitat bàsica de la cromatina eucariòtica és el nucleosoma, integrat per quatre proteïnes histones que conformen un octàmer al voltant del qual es disposa la doble hèlix d’ADN en dos girs “superhelicals”. Aquest tret estructural fa que l’ADN quedi especialment protegit. Un sistema complex de control d’accés permet a la maquinària proteica responsable de funcions com la transcripció, la replicació o la reparació d’accedir a l’ADN. Modificacions post-transcripcionals de les histones, com la metilació, constitueixen un element central de la regulació epigenètica.
La història de la recerca en histones arrenca en el 1884, quan Albrecht Kossel reportà l’aïllament d’un component semblant a la peptona i extractable amb àcids dels nuclis d’eritròcits d’oca. Kossel anomenà a aquest component “histona” sense aclarir si ho feia pels termes grecs ἵστημι o ἱστός, però sí assenyalant que sospitava que in vivo aquesta histona es trobaria associada a l’àcid nucleic en els nuclis intactes d’eritròcit. El grup de recerca de James F. Bonner, en els anys 1960, atenent al fet que hom havia descobert histones molt abundants i semblants entre els teixits i organismes més diversos, estudià aquesta proteïna i determinà la seva importància en la regulació dels àcids nucleics, dels quals ja se sabia que eren la molècula de l’herència biològica. La biologia estructural descrigué el ‘plec d’histona’ com un motiu integrat per tres hèlix alfa connectades per dos llaços curts. Aquest ‘plec d’histona’ permet la dimerització de dues molècules d’histona com qui es dóna la mà, tocant el cap d’una amb la cua de l’altra.
Si d’entrada hom els assignava un rol essencialment estructural, de plegament d’ADN, com més anava més evident era el seu paper funcional en la regulació de l’expressió gènica i del cicle cel·lular. Durant força temps hom pensà que les histones eren exclusives dels organismes eucariòtics, però eventualment hom detectà la presència d’histones relativament senzilles (consisteix en un únic plec d’histona) en arqueons. Així hom conclogué que en arqueons hi ha una organització de la cromatina dependent d’aquestes histones minimalistes i d’altres proteïnes associades al nucleoide (NAPs). Aquesta troballa encaixava amb la idea que els arqueons, com a organismes procariòtics, són un pont evolutiu entre els bacteris i els eucariotes. En alguns arqueons les histones són les proteïnes cel·lulars més abundants. En Thermococcus kodakarensis la presència d’histones és imprescindible per a la viabilitat. Els dímers d’histones d’aquests arqueons són envoltats per ADN en contactar-hi a través de tres solcs consecutius de la doble hèlix, és a dir d’una manera semblant a la del nucleosoma eucariòtic. Existeixen, però, importants diferències. Les histones d’eucariotes són heterodímers obligats. En canvis les histones HTkA/B (T. kodakarensis) i HMfA/B (de Methanothermus fervidus) poden formar tant heterodímers com homodímers. Si els heterodímers d’histones eucariotes s’integren en octàmers ben definits, els dímers d’histones arqueanes oligomeritzen en hipernucleosomes de mida variable.
Quan diem que la cèl·lula bacteriana és una cèl·lula procariota, ens referim al fet que el nucli (o nucleoide) cel·lular no es troba separat per una membrana de la resta del citoplasma. L’ADN bacterià, però, o es troba nuu, sinó que en la seva organització participen una sèrie de proteïnes bàsiques NAPs amb capacitat d’unió a l’àcid nucleic. Entre aquestes NAPs hi ha HU i d’altres. Algunes NAPs són exclusives de llinatges bacterians concrets, i d’altres, com la citada HU, són més generals. Ara bé, la deleció de la proteïna HU, que és la NAP més abundant d’Escherichia coli no implica letalitat. Aquesta troballa reforçà la idea d’un rol menor de les NAPs en l’organització del material genètic bacterià, i l’assumpció que no hi havia histones entre els bacteris.
En ampliar-se les bases de dades genòmiques de bacteris, les recerques bioinformàtiques posaren de manifest la presència de possibles dominis de ‘plegament d’histona’ en una fracció petita però variada de llinatges bacterians.
Hocher et al. presenten en aquest estudi resultats que indiquen que en el bacteri Bdellovibrio bacteriovorus les histones són components majoritaris i essencials del nucleoide. In vitro, aquestes histones recobreixen l’ADN linear constituint un filament de nucleohistona. Si entre els eucariotes és l’ADN el que encercla l’octàmer d’histones, en B. bacteriovorus és la histona la que embolcalla la doble hèlix d’ADN.
Una recerca de proteïnes amb plegament d’histona
Hocher parteixen dels dominis de plegament d’histona PF00125, PF00808 i PF09123 per fer una recerca bioinformàtica sobre una base de dades amb 18.343 genomes bacterians del NCBI. Descartaren els resultats corresponents a proteïnes majors de 200 aminoàcids.
Els cultius bacterians analitzats
La recerca assenyalà com a objectiu a B. bacteriovorus HD100. Aquest bacteri és cultivat en plaques que contenen E. coli S17-1, de les quals s’alimenta.
B. bacteriovorus HID13 fou seleccionat pel fet que se’l pot cultivar en el laboratori sense necessita d’un bacteri hoste.
Un altre objectiu fou L. interrogants, que és cultiva en un medi EMJH a 30°C.
Dels cultius bacterians es feien extraccions de proteïna total. També es feien extraccions a partir d’un enriquiment previ de nucleoides cel·lulars. Els extractes de proteïna eren processats per analitzar-los per cromatografia líquida acoblada a espectrometria de masses (LC-MS/MS).
De cultius bacterians també es feia extracció de l’ARN total, que era ulteriorment analitzat en una seqüenciador.
El marcatge amb fluorescència de Bd0055
Mitjançant enginyeria genètica es fusionà el fluoròfor mCherry/mCitrine a l’extrem C- termina de la proteïna Bd0055. Això permetia fer un seguiment d’aquesta histona en microscopi d’epifluorescència.
L’expressió de Bd0055 en E. coli
El gen Bd0055 fou introduït en un vector d’expressió d’E. coli. Així s’aconseguien quantitats d’aquesta proteïna inassolibles en cultius de B. bacteriovorus. Aquesta quantitat de proteïna serví per als estudis de cristal·lografia, i per als experiments d’unió a ADN i de dinàmica molecular.
Mutagènesi dirigida
Hocher et al. intentaren generar mutants de B. bacteriovorus que tinguessin el gen bd0055 delecionat. Quelcom semblant intentaren també en L. interrogans per al gen LA_2458.
Les histones són presents en diversos clades bacterians
En els 18.343 genomes bacterians investigats, Hocher et al. troben 416 proteïnes que contenen un domini de plegament d’histona. No arriba al 2% els genomes bacterians que contenen un domini de plegament d’histones. En canvi, el 93% dels genomes bacterians contenen la proteïna HU. Això ja indica que no hem de pensar pas que les proteïnes de tipus histona siguin gaire abundants o escampats entre els bacteris.
La distribució filogenètica d’histones entre el domini bacterià és indicativa d’una transmissió vertical en certs llinatges i de transmissió horitzontal en uns altres. Hi ha dos clades que s’hi destaquen en la persistència d’histones: Bdellovibrionota i Leptospirales.
Bdellovibrio bacteriovorus HD100 és un predador o paràsit bacterià amb un estil de vida bifàsic: 1) fase de vida lliure; 2) fase invasiva sobre el periplasma de bacteris gramnegatius com E. coli. No és estrany que des de la seva descoberta, en el 1962, hom hagi somniat en utilitzar aquesta “sangonera de bacteris” per combatre infeccions bacterianes en humans i animals. Hocher et al. mostren que en el genoma hi hauria dos gens de possibles histones: Bd0055 i Bd3044. Les dades transcriptòmiques prèvies assenyalaven una alta expressió de Bd0055 particularment durant la replicació activa dins del periplasma de l’hoste. Per això, Hocher et al. decidiren centrar-se en aquesta proteïna.
Hocher et al. remarquen en les imatges de microscopi de fluorescència amb Bd0055 marcada, que aquesta proteïna és absent dels pols cel·lulars, i tampoc no és secretada. La tinció Hoechst indica que es localitza en el nucleoide. La proteïna Bd0055 es troba enriquida 29 vegades en la fracció de nucleoides. En aquesta fracció Bd0055 és la més abundant de les NAPs, superant HU i Dps.
La cristal·lografia indica que Bd0055 forma dímers en els que els monòmers es connecten amb el cap de l’un tocant la cua de l’altre.
En els assaigs d’unió a fragments de 147 bp d’ADN, la proteïna Bd0055 s’uneix sense formar oligòmers estables. L’anàlisi cristal·logràfica de la unió de la proteïna Bd0055 amb fragments d’ADN de 35 bp indica un contacte a través d’un solc menor de la doble hèlix. La doble hèlix, però no es plega al voltant de la proteïna Bd0055, sinó que és la proteïna Bd0055 la que acaba per encerclar l’ADN format un filament de nucleohistona amb una ratio d’un dímer de Bd0055 per cada 2,5 bp d’ADN.
La introducció de mutacions A48H, S45F i I61L a la proteïna Bd0055 permet que aquesta segueixi un comportament més semblant a les histones arqueanes i eucariòtiques. Aquest comportament seria degut a la formació de tetràmers de Bd0055.
L’anàlisi de mutants per deleció del gen bd0055 indica que la proteïna Bd0055 és essencial per al creixement de B. bacteriovorus tant en una soca depredadora com HD100 com en una de vida lliure com HID13.
L’altra histona investigada per Hocher et al. és A0A2H1XGH2, codificada pel gen LA_2458 de Leptospira interrogans. Aquesta espiroqueta és l’agent causal de la leptospirosi que en el nostre entorn afecta sobretot gossos. Aquesta histona també es troba enriquida en el nucleoide, i el gen LA_2458 resulta essencial per a la viabilitat del bacteri.
Hocher et al., doncs, mostren dos exemples de bacteris on histones participen en l’organització de la cromatina. La histona Bd0055 mostra in vitro una interacció amb l’ADN que hom no coneixia entre les histones d’eucariotes i arqueons: la de formar-hi una fibra densa que encapsula la doble hèlix. Hocher et al. aporten indicis en forma de tomogrames de B. bacteriovorus i L. interrogans en els que semblaria que aquestes fibres s’arriben a formar també in vivo. Hipotetitzen que aquesta compactació en fibres de l’ADN ajudaria a la contracció del nucleoide de B. bacteriovorus en la fase prèvia a l’entrada de la cèl·lula depredada o parasitada. Potser la histona de L. interrogans fa un paper semblant en la compactació del nucleoide necessària per al manteniment de la forma filamentosa pròpia d’una espiroqueta. En tot cas tant aquesta com l’altra hipòtesi requeriran estudis addicionals sobre aquests dos bacteris.
En tot cas hem de corregir una idea encara massa arrelada. On dèiem que “en els bacteris no hi ha histones”, ara haurem de dir “en alguns bacteris hi ha histones”.

Electromicrografia on s'observen dues cèl·lules de l'espiroqueta "Leptospira interrogans"
Lligams:
- Histone-organized chromatin in bacteria. Antoine Hocher, Shawn P. Laursen, Paul Radford, Jess Tyson, Carey Lambert, Kathryn M Stevens, Mathieu Picardeau, R. Elizabeth Sockett, Karolin Luger, Tobias Warnecke. https://doi.org/10.1101/2023.01.26.525422