Biofísica quàntica: En la funció de l’àcid desoxiribonucleic (ADN) com a molècula portadora de la informació genètica resulta essencial la minimització de les mutacions que afectin la seqüència genètica de mononucleòtids. En la molècula de doble cadena (doble hèlix) d’ADN, les dues cadenes polinucleotídiques es vinculen a través de ponts d’hidrogen que depenen de la seqüència nucleotídica. La transferència de protons a través d’aquests ponts d’hidrogen pot induir un fenomen de tautomerització que condueixi a una eventual mutació puntual en la seqüència nucleotídica. Louie Slocombe, Marco Sacchi i Jim Al-Khalili proposen en un article a Communication Physics una anàlisi teòrica dels ponts d’hidrogen que es formen entre els nucleòtids de guanina i de citosina de dues cadenes complementàries d’ADN. Aquesta anàlisi inclou un model acurat de l’estructura del parell de bases nitrogenades, la dinàmica quàntica del protó del pont d’hidrogen i la influència decoherent i dissipativa de l’ambient cel·lular. La contribució de l’efecte túnel quàntic a la taxa de transferència protònica és diverses ordres de magnitud superior al salt de la barrera de física clàssica. Com a conseqüència de l’efecte túnel hi ha una interconversió entre les formes canònica i tautomèrica dels ponts G-C fins que s’arriba a un equilibri tèrmic. La probabilitat d’ocupació tautomèrica que calculen Slocombe et al. és de 1,73·10-4, força coincident amb les taxes de mutació.
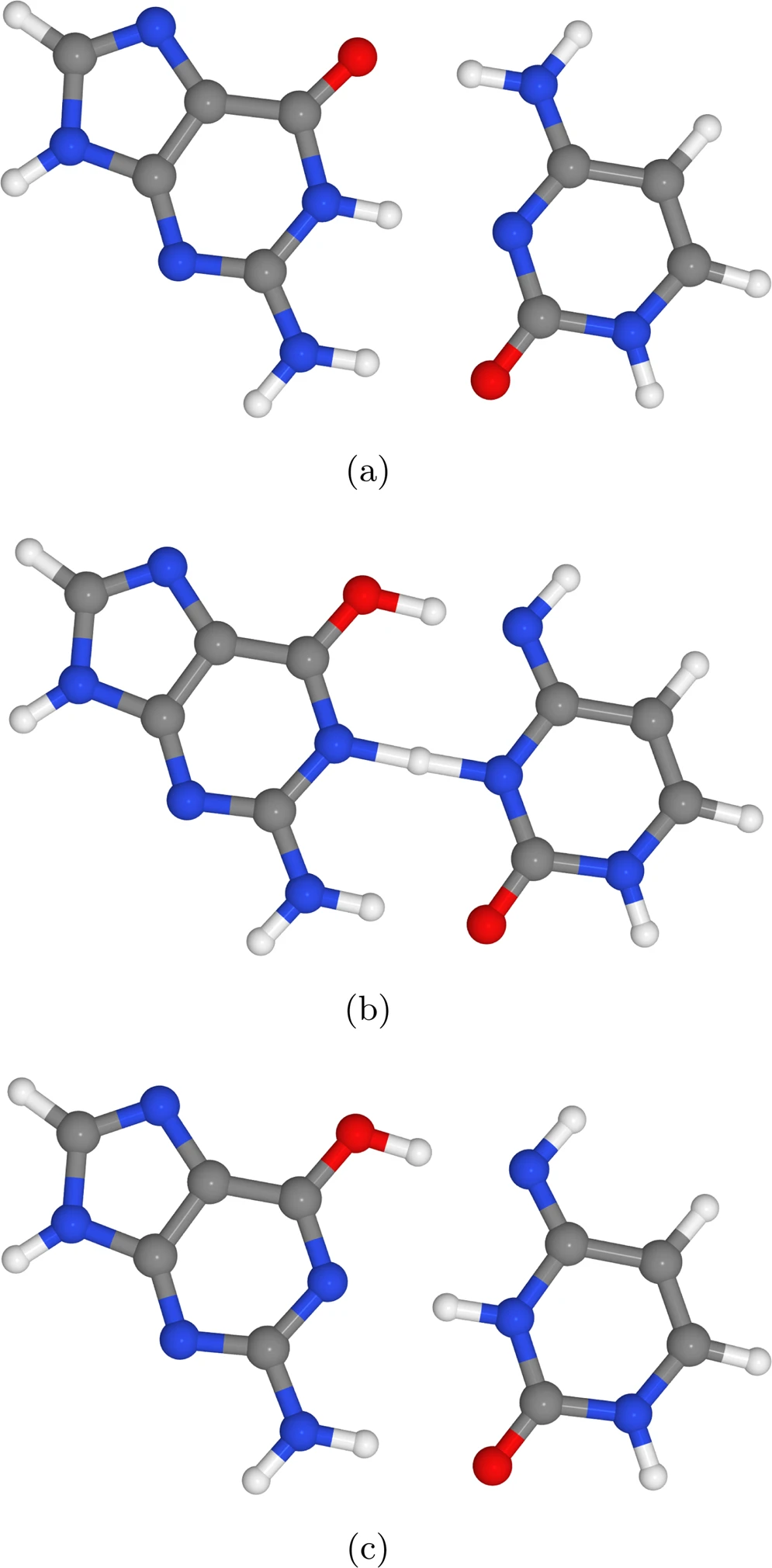
L’ADN consisteix en un polímer de mononucleòtids. Cada mononucleòtid es caracteritza per la presència d’una base nitrogenada. Les quatre bases nitrogenades de l’ADN són la guanina (G), la citosina (C), l’adenina (A) i la timina (T). La complementarietat entre G i C, i A i T, és la que permet la formació de la doble hèlix característica de l’ADN genòmic. Slocombe et al. s’ocupen concretament de la formació de ponts d’hidrogen entre G i C
Una anàlisi teòrica dels ponts d’hidrogen G-C de la doble hèlix d’ADN
Sacchi i Al-Khalili conceberen i dissenyaren aquesta recerca. Sacchi és membre del Department of Chemistry de la University of Surrey, i Al-Khalili del Department of Physics del mateix centre. Slocombe, que fa la tesi doctoral en aquesta universitat, construí l’aparell computacional. Els tres autors tenen paraules d’agraïment pels suggeriment de Johnjoe McFadden, Max Winokan i Antonio Pantelias.
El model de la doble hèlix de l’ADN fou postulat en el 1953 per Watson & Crick, que ja llavors suggeriren que la tautomerització de parells de bases podia trobar-se a la base de mutacions permanents. En el model de Watson & Crick, entre les guanines (G) i les citosines (C) es formen tres ponts d’hidrogen, i entre les adenines (A) i les timines (T), dos. La doble transferència de protons a través d’aquests ponts pot conduir a una tautomerització.
Watson & Crick únicament contemplaven els fenòmens de la física clàssica. En els darrers anys, però, ha crescut l’interès per la participació de fenòmens quàntics en l’estructura de l’ADN. L’efecte túnel dels protons dels ponts d’hidrogen ha estat postulat com a contribuïdor en les relacions entre les dues cadenes de l’ADN bicatenari.
La tautomerització resulta de la transferència de protons del pont d’hidrogen des de la base d’una cadena a la complementària: la posició de totes dues cadenes queda alterada. En la replicació de l’ADN aquesta tautomerització pot persistir després de l’obertura de la doble hèlix per l’helicasa i heretar-se en les cadenes de nova síntesi.
En el cas d’un parell G-C, la tautomerització es representa com a G*-C*. Si la tautomerització passa els filtres de l’helicasa i del replisoma, es pot formar un parell G*-T, que doni eventualment com a resultat la mutació G>A.
Hom pot dir que la transferència doble de protons és un fenomen massa efímer per a sobreviure al procés de replicació. L’acció de l’helicasa, per exemple, té lloc a una escala de nanosegons. Slocombe et al., però, assenyalen que allò rellevant no és la durada de l’estat tautomèric, sinó la fracció de parells G-C que es trobin en estat tautomèric en el moment de la replicació de l’ADN.
La tautomerització dependria de la transferència d’un protó a través de la barrera entre les dues cadenes, acompanyada del moviment de l’altre protó en la direcció oposada que preserva l’electroneutralitat del sistema.
Com tota reacció química reversible, G-C > G*-C* té una constant d’equilibri (Keq) que és la ratio entre la concentració de G*-C* i la concentració de G-C. Slocombe et al. afegeixen, a més, un coeficient d’activitat quàntica Γqm, que té en compte l’efecte túnel reforçat per la temperatura, l’energia de punt zero, la decoherència i la dissipació.
El càlcul de Keq ha estat estimat teòricament (a través de la diferència en l’energia lliure de Gibbs de les formes canòniques i tautomèriques de G-C) i també experimentalment. Slocombe et al. ho complementen amb un càlcul de mecànica quàntica basat en un sistema quàntic obert. Aquest recàlcul augmenta la taxa de tautomerització de valors clàssics de 10-8 a 10-4.
En comparació amb G*-C*, A*-T* és extremadament inestable, i per això Slocombe et al. pensen que els parells A-T tenen una importància menor en la mutagènesi espontània. En canvi, els efectes quàntics en els parells G-C tindrien un caràcter protagònic, i serien un exemple de genètica quàntica.
Un sistema quàntic obert
Slocombe et al. descriuen el paisatge d’energia potencial del doble pont d’hidrogen entre G i C atenent a l’ambient cel·lular. La ΔE de 0,435 eV es troba entre una barrera alta de 0,705 eV i una barrera reversa de 0,270 eV:

En el sistema quàntic obert es contempla un bany tèrmic infinit d’oscil·ladors harmònics. La dinàmica interna de Schrödinger és complementada amb els fenòmens de dissipació i de decoherència.
Si pensem en el context de la genètica humana, la temperatura rellevant d’aquest bany és de 300 K. L’ambient té natura aquosa, de manera que s’assum un espectre de col·lisió en el rang de 3300-3900 cm-1.
El factor túnel tindria una contribució cabdal en la taxa de reacció i en la vida mitjana de les formes canònica i tautomèrica dels parells G-C. La transferència de protons es realitza a una escala temporal de pocs centenars de femtosegons (10-13 s). A efectes pràctics, és com si els protons poblessin simultàniament tots dos estats, de manera que el parell G-C transita contínuament entre les formes canònica i tautomèrica.
Mentre que la transferència de protons en el pont d’hidrogen es realitza en períodes de 10 femtosegons a 100 nanosegons, l’helicasa triga molt més de 100 nanosegons en separar les dues cadenes de l’ADN. La taxa de tautomerització corregida quànticament permet estimar la fracció de parells G-C que entren en el replisoma tautomeritzats.
Física quàntica en la mutació espontània de l’ADN
Segons el model de Slocombe et al. la probabilitat que un parell G-C entri en el replisoma en la forma tautomèrica és d 1,73·10-4. La probabilitat és quatre ordres superior a l’estimada amb models clàssics.
Lligams:
- An open quantum systems approach to proton tunnelling in DNA. Louie Slocombe, Marco Sacchi & Jim Al-Khalili. Communication Physics 5: 109 (2022).
Cap comentari:
Publica un comentari a l'entrada