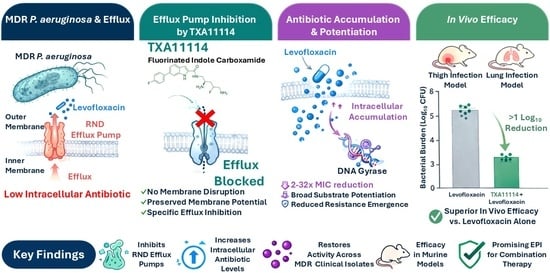
Antropologia: El Grup de Treball Waka Kotahi Tangata Whenua nasqué amb la intenció de garantir la representació de marae i hapu locals en el projecte de construcció de la Via Ràpida de Waikato en la zona de Taupiri Maunga, muntanya sagrada per als Waikato-Tainui. En el curs de la construcció es descobrí accidentalment un enterrament de tūpuna (ancestres). L’antròpologa Rebecca L. Kinaston és l’autora primera i corresponsal d’un article a Nature Communications on s’indaga en l’alimentació d’aquests ancestres del segle XVIII. Val a dir que les històries orals maori, els relats etno-històrics i dades arqueològiques ja indicaven a que l’horticultura del kūmara (Ipomoea batatas) i el taro (Colocasia esculenta) foren claus en el creixement demogràfic i el canvi cultural d’aquest període. Kinaston et al. han investigat la dieta, el lloc de residència i el sexe cromosòmic d’aquestes kōiwi tangata (restes humanes). Les dades arqueològiques assenyalen que ja des del segle XVI a la regió hi havia una horticultura intensiva. Les restes humanes investigades remuntarien al període 250-170 abans del Present, i es correspondrien a un enterrament secundari compartit en una fossa excavada durant el Període Tradicional de la història Māori, en un ric context artístic, arquitectònic, cosmològic i genealògica (whakapapa). Les anàlisis isotòpics i de pèptids d’esmalt indiquen que els set tūpuna seguien una dieta basada principalment en aliments vegetals. Dos infants (cromosòmicament mascle i femella) eren probablement locals i foren deslletats amb aliments vegetals en el segon o tercer any de vida. Així doncs, les investigacions refermen la idea de la centralitat de l’horticultura a Waikato durant el Període Tradicional, amb una dieta predominantment vegetal.

El riu Waikato al seu pas per Hamilton
El Període Tradicional Maori
Les dades arqueològiques situen el primer assentament humà d’Aotearoa a mitjan segle XIII. Aquests habitants procedien de la Polinèsia Oriental. Algunes històries maorís conviden a pensar en un poblament més antic, però coincideixen en contrastar la terra tropical d’origen d’aquests pobladors, Hawaiki, amb la climatologia temperada d’Aotearoa.
Els jaciments del Període Inicial (1250-1450 d.C.) d’Aotearoa assenyalen que els pobladors depenien d’una dieta variada de recursos animals marins i terrestres. Eren poblacions nòmades, que realitzaven circumnavegacions al voltant de les dues illes principals. També introduïren collites hortícoles procedents de la Polinèsia Oriental. Transformaren el paisatge a gran escala. La caça especialitzada en grans mamífers marins i aus terrestres (moa) adquirí gran rellevància.
En el Període Mitjà o Transicional (1450-1650 d.C.) i, encara més en el Període Tradicional (1650-1769 d.C.), hi hagué una tendència cap a un assentament més regionalitzat. A Te Ika-a-Maui (l’illa del Nord) hi hagué una intensificació hortícola més marcada que a Te Waipounamu (l’illa del Sud). Tot i amb tot, les activitats recol·lectores (mahinga kai) continuaren.
A la Conca del riu Waikato, al voltant de la ciutat de Hamilton (Kirikiriroa), les històries orals, els relats etno-històrics i el registre arqueològic ha permès definir el Complex Hortícola de Waikato. Des del segle XVI s’hi hauria fet una horticultura intensiva, gràcies als sòls al·lofànics derivats de tefra, rics en carboni orgànic i amb un bon drenatge i aireig.
Els principals tubercles cultivats pels maoris eren la kūmara i, en menor mesura, el taro i l’uhwikāho (Dioscorea alata). Tots ells foren introduïts a Aotearoa des de la Polinèsia Oriental, possiblement en diferents etapes. La base de la intensificació hortícola foren els aclariments de boscos ja iniciats des del 1250 d.C. A Waikato els horts eren establerts després d’abatre arbres i cremar-hi les soques. Els sòls eren adobats amb material lític (sorra i grava), que era extret del subsòl.
Els jaciments arqueològics assenyalen aquesta activitat hortícola, alhora que mostren una absència de restes faunístiques (de fauna marina, fluvial o terrestre).
Kinaston et al. presenten en aquest treball una investigació sobre les vides de set tūpuna que haurien viscut entre el 1700 i el 1780, i que foren enterrats en una fossa. En primer lloc fan una descripció del ritual de soterrament d’aquestes kōiwi tangata, i seguidament un estudi osteològic i una anàlisi d’esmalt dentari per fracció isotòpica i peptídica. També fan una determinació del sexe cromosòmic. Les dades permeten deduir pràctiques de deslletament i lloc de residència durant la infantesa.
Set kōiwi
Kinaston et al. presenten resultats de la distribució isotòpica de la col·làgena òssia de set kōiwi. Tan sols de dos individus (Kōiwi 2 i Kōiwi 4) presentaven primers molars permanents, i és d’ells que es publiquen dades de pèptids d’esmalt, de fracció isotòpica (δ13C i δ15N) de dentina i de fracció isotòpica (δ13C, 87Sr/86Sr) d’esmalt.
Pèptids d’esmalt
Tant de Kōiwi 2 com de Kōiwi 4 es recuperaren pèptids d’amelogenina de les mostres de dents. Kōiwi 4 presenta els pèptids AMEL-X i AMEL-Y, indicatius de sexe cromosòmic masculí. Kōiwi 2 tan sols presenta pèptids AMEL-X, i se’n dedueix un sexe cromosòmic femení.
Les fraccions isotòpiques δ13C i δ15N de col·làgena òssia
De tots set individus es van poder prendre mostres de col·làgena òssia de prou qualitat (amb un %N d’entre 11-16%; amb un %C > 30%; i una ratio C:N d’entre 2,9 i 3,6).
Kinaston et al. admeten que es tracta d’una mostra petita, de 3 adults i de 4 no-adults. Aventuren la dada que els adults tenien un valor de δ15N un 0,7‰ superior als no-adults, entre que el valor de δ13C seria més semblant. Els no-adults presentaven una edat osteològica d’entre 5 i 15 anys.
Les fraccions isotòpiques δ13C i δ15N de dentina
Les mostres de micropunxada de dentina segueixen un criteri de qualitat semblant al de la col·làgena, per bé que la ratio C:N s’hauria de situar entre 3,1 i 3,6.
Per als dos individus analitzats, Kōiwi 2 i Kōiwi 4 es fa una comparació dels valors de δ13C de la dentina coronal de la junció cemento-esmalt, i del carbonat de l’esmalt. Això permet una estimació de la contribució a la dieta de plantes C3 i de plantes C4/aliments marins durant el període d’infantesa (1,9-3,5 anys d’edat). Kōiwi 2 i Kōiwi 4 seguien una dieta basada en plantes terrestres C3.
La fracció isotòpica d’estronci de l’esmalt dentari.
La ratio 87Sr/86Sr és informativa d’origen geogràfic si hom atén al mapa isotòpic d’estronci d’Aotearoa. En efecte, la ratio depèn del lloc de residència en el període d’infantesa (1,9-3,5 anys d’edat). Kōiwi 2 i Kōiwi 4 haurien crescut probablement a Tamahere, o potser als Llacs de Rotorua. No obstant, Kinaston et al. pensen que l’edat primerenca de la mort (7-9 anys en el cas de Kōiwi 2; i 9-15 anys en el cas de Kōiwi 4) s’adiu amb un origen local a Tamahere.
Localitat i dieta vegetal
Les dades isotòpiques òssies i dentals dels set tūpuna de Tamahere indiquen una dieta basada principalment en vegetals. Cal pensar en una dieta basada en plantes C3 com la kūmara i, en menor mesura, taro i uwhikāho. Kinaston et al. descarten altres fonts vegetals atenent a un desgast dental no pas sever. Ara bé, aquesta dieta vegetal es tradueix en una taxa elevada de caries.
Els valors isotòpics de col·làgena òssia i de dentina representen principalment la proteïna de la dieta, ja que són els aminoàcids procedents dels aliments els qui es vehiculen preferencialment per a la síntesi de teixit. Les dades dels individus de Tamahere reflecteixen una relativa absència d’aliments rics en proteïna com carn o peix. Efectivament els valors baixos en la col·làgena de δ13C i δ15N indiquen una font proteica de baix nivell tròfic, concretament de plantes terrestre que fan la fotosíntesi C3. El consum de proteïna animal devia ser esporàdic, potser aus silvestres, o bé kurī (Canis familiaris) o kīore (Rattus exulans) alimentats amb kumara i altres tubercles.
Mig mil·lenni després de la colonització inicial d’Aotearoa, els maoris de Waikato ja havien desenvolupat plantacions prou productives com per collir emmagatzemar prou tubercles per a fer-ne el principal aliment durant tot l’any, per bé que el cultiu de la kūmara és estacional. A la regió hi hauria hagut uns 500 horts, i 200 llocs fortificats (pā). Un subproducte de l’activitat hortícola foren milers de fosses excavades prop dels rius Waikato i Waipā.
Aquesta dieta vegetal impactava en el deslletament dels infants. Kōiwi 2 mostra la caiguda en el valor de δ15N de la dentina entre el naixement i els 2,5 anys d’edat. Kōiwi 4 hauria seguit un deslletament complet cap als 3,2 anys d’edat.
L’enterrament en fossa on es trobaren les restes d’aquests individus era secundari i col·lectiu. La tradició oral explica aquests enterraments secundaris amb l’objectiu d’evitar que els enemics s’apoderessin de les restes d’enterraments primaris per robar-ne la força vital (mana). Les restes humanes analitzades no mostres signes de violència però sí marques de tall postmortem.
Lligams:
- Horticultural intensification and plant-based diets of 18th century CE Waikato Māori in Aotearoa New Zealand. Rebecca L. Kinaston, Sian Keith, Beatrice Hudson, Jonny Geber, Torsten Kleffmann, Claudine Stirling, Malcolm Reid, David Barr, Robyn Kramer, Emma Sudron & Waka Kotahi Tangata Whenua Working Group. Nature Communications 17: 3040 (2026).